
Eiconaxius acutifrons Bate, 1888
|
publication ID |
https://doi.org/10.5281/zenodo.214595 |
|
DOI |
https://doi.org/10.5281/zenodo.6181685 |
|
persistent identifier |
https://treatment.plazi.org/id/03AD87EB-FFF2-5427-FF0D-63BCFCA67688 |
|
treatment provided by |
Plazi (2016-04-12 22:29:05, last updated 2024-11-28 14:29:16) |
|
scientific name |
Eiconaxius acutifrons Bate, 1888 |
| status |
|
Eiconaxius acutifrons Bate, 1888 View in CoL
( Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Eiconaxius acutifrons Bate, 1888: 40 View in CoL , fig. 4, pl. 5, fig. 2d, g, h, i, k, l; Sakai & de Saint Laurent 1989: 16 (in part); Kensley 1996: 475 (list); Sakai, 2011: 270.
Axius (Eiconaxius) acutifrons: Borradaile 1903: 538 (list).
Iconaxius acutifrons . –– Balss 1925: 209 (list; in part).
Not Axius acutifrons . –– Faxon 1895: 103, pl. 28, fig. 2. = Eiconaxius albatrossae Kensley, 1996 View in CoL .
Not Axius (Eiconaxius) acutifrons . –– De Man 1925: 37, pl. 3, fig. 5–5e. = Eiconaxius indonesicus Sakai, 2011 View in CoL .
Not Axius acutifrons . –– Wicksten, 1982: 246, fig. 1; 1989: 313 (list). = Eiconaxius baja Kensley, 1996 View in CoL . Not Eiconaxius acutifrons View in CoL . –– Hendrickx 1995: 157 (list).
Material examined. Lectotype (designated herein): male (cl 5.0 mm), Banda Sea, Indonesia, H. M. S. Challenger, stn 194A, 04°31.0’S, 129°57.20’E, 658 m, volcanic mud, 29 September 1874, NHM 1888.22.
Paralectotype: 1 ovigerous female (cl 6.9 mm), same data as lectotype.
Non-type: 1 male (cl 4.4 mm), Myojin Knoll, northern Izu Islands, 32°05.952’N, 139°52.050’E, 1074 m, RV Kairei, KR10-E02 cruise, ROV Kaiko 7000II, dive 486, 17 September 2010, JAMSTEC 083387; 2 males (cl 3.6, 3.9 mm), same data, JAMSTEC 83398, 83713.
Description. Lectotype (male). Rostrum ( Fig. 1 View FIGURE 1 A, B) narrowly triangular in dorsal view (about 1.9 times longer than basal width), apically narrowly rounded, directed forwards, reaching midlength of second segment of antennular peduncle; lateral margins each with 3 tiny obtuse teeth; apex upturned. Carapace ( Fig. 1 View FIGURE 1 A, B) with gastric region slightly convex, slightly sloping down to rostrum; cervical groove absent; median carina low, entire, not markedly broadened posteriorly, nor bifurcated; submedian carina absent; lateral carinae diverging posteriorly, reaching to about anterior one-third of carapace length.
Abdomen ( Fig. 1 View FIGURE 1 C) slightly depressed dorsoventrally. First abdominal somite with pleuron poorly defined ventrally. Second and third pleura produced posteroventrally into sharp tooth; fourth pleuron produced into subacute tooth ventrally, with tiny sharp tooth anteroventrally; fifth pleuron generally rounded, with tiny tooth on ventral margin. Sixth somite subequal to fifth somite, posterodorsal margin weakly denticulate; pleuron bluntly triangular; posterolateral process acuminate. Telson ( Fig. 2 View FIGURE 2 A) with greatest width at anterior 0.2; lateral margin bearing 6 (left) or 5 (right) tiny teeth, posteriormost tooth located at posterolateral angle; no movable spine at posterolateral angle; posterior margin nearly straight, bearing small median tooth.
Eye ( Fig. 1 View FIGURE 1 A, B) not reaching midlength of rostrum, subglobose; cornea not faceted. Antennular peduncle ( Fig. 1 View FIGURE 1 A, B) moderately stout; distal two segments subequal in length to first segment. Antennal peduncle ( Fig. 1 View FIGURE 1 A, B) moderately stout; distolateral prolongation of second segment acute, slightly overreaching distal margin of third segment of antennular peduncle; third segment with small tooth at ventromesial distal angle; antennal acicle large, acuminate, reaching beyond distal margin of fifth segment of antennal peduncle.
Third maxilliped similar to that of paralectotype.
Major (right) cheliped ( Fig. 1 View FIGURE 1 D, E) massive. Ischium with 3 tiny to minute spiniform denticles on ventral margin; dorsal margin unarmed. Merus strongly compressed laterally; dorsal margin arcuate, bearing 3 minute denticles in distal half (distalmost denticle terminal in position), otherwise nearly smooth; ventromesial margin sharply carinate, with 2 tiny subterminal denticles followed by 5 minute denticles; laterodistal projection rounded. Carpus cup-shaped, much higher than long, ventral angle with minute denticle. Chela slightly longer than carapace (including rostrum). Palm 1.1 times longer than high; dorsal margin sharply carinate, terminating distally in small acute tooth, further bearing 2 tiny denticles in distal 0.4; lateral surface convex, glabrous, with some scattered low tubercles in distal 0.3; ventrolateral carina sharp, extending nearly to tip of fixed finger, margin upturned; mesial surface also with some scattered low tubercles in anterior 0.3. Fixed finger nearly straight but distally slightly upturned; opposable margin with 1 prominent triangular tooth at midlength and row of several minute denticles in distal half, and with small, shallow hiatus proximally. Dactylus 0.9 times as long as palm, terminating in slightly curved calcareous claw, dorsal margin sharply carinate, unarmed; lateral surface slightly elevated along midline, shallowly sulcate along dorsal carina; opposable margin with 1 large blunt cusp proximally, otherwise unarmed, no prominent notch or excavation distal to cusp.
Minor (left) cheliped ( Fig. 1 View FIGURE 1 F) subequal in length to major cheliped. Ischium with 2 tiny denticles on dorsal and ventral margins respectively. Merus strongly compressed laterally; dorsal margin generally convex, sharply carinate, bearing 3 minute denticles distal to midlength; ventromesial margin sharply carinate, with 2 minute denticles and 1 small subterminal tooth; laterodistal projection subacute. Carpus cup shaped, much higher than long, ventral angle unarmed. Chela subequal in length to carapace (including rostrum); ventral margin gently sinuous. Palm slightly becoming higher distally, nearly as long as high; dorsal margin sharply carinate, bearing 1 minute denticle subdistally and 1 tiny spiniform tooth distally; lateral surface convex, smooth; ventrolateral carina sharp, extending nearly to tip of fixed finger; mesial surface smooth. Fingers forming deep excavation defined by sharp longitudinal carina on mesial side. Fixed finger almost straight but distally slightly upturned, with row of small, triangular or rounded teeth over entire length; dactylar articulating margin with 2-spined cusp laterally; lateral face flanked by blunt upper ridge along cutting edge and ventrolateral carina concave. Dactylus about 1.5 times as long as palm, terminating in slightly curved calcareous claw, dorsal margin sharply carinate; lateral surface with blunt longitudinal carina on midline; cutting edge unarmed.
Second to fourth pereopods of similar length, moderately stout. Second pereopod ( Fig. 2 View FIGURE 2 B) unarmed on ischium to carpus; chela about 1.4 times longer than carpus, with scattered tufts of setae; fixed finger deflexed, with row of minute corneous spinules on cutting edge; dactylus about 0.5 times as long as palm, setose, also with row of minute corneous spinules on cutting edge. Third pereopod ( Fig. 2 View FIGURE 2 C) unarmed on ischium to carpus; propodus about 1.5 times longer than carpus, with 9 sets of spinules on lateral surface ventrally (each set consisting of a single spinule or paired spinules) and single spinule at ventrodistal margin; dactylus strongly compressed laterally, suboval, terminating in clearly demarcated claw, with about 9 accessory spinules on flexor margin and 2 submarginal spinules proximally. Fourth pereopod ( Fig. 2 View FIGURE 2 D) similar to third pereopod; propodus with 5 sets of spinules (each set consisting of 2 or 3 transverse row of spinules); dactylus 10 accessory spinules on flexor margin. Fifth pereopods shorter than preceding ones; propodus with grooming apparatus consisting of oblique row of stiff setae and slender spinules on distal part; dactylus with 10 accessory spinules on flexor margin.
First pleopod absent. Second pleopods not preserved. Third to fifth pleopods slender, biramous, each with appendix interna. Uropodal exopod ( Fig. 2 View FIGURE 2 E) with lateral margin serrated with 15 minute to tiny acute teeth on lateral to posterior margin (posteriormost tooth strongest), mesial margin regularly convex; endopod with lateral margin serrated with 8 tiny acute teeth (posteriormost tooth strongest).
Paralectotype (female). Generally similar to lectotype except for sexual characters. Rostrum ( Fig. 3 View FIGURE 3 A) relatively broader than in lectotype (about 1.4 times longer than wide); lateral margins only faintly denticulate. Abdomen ( Fig. 3 View FIGURE 3 C) proportionally broader than in males. Telson with 4 tiny teeth on lateral margin. Third maxilliped ( Fig. 3 View FIGURE 3 D) moderately slender for genus; basis with 1 tiny spine on ventral margin distally; ischium bearing row of tiny denticles on mesial crest (crista dentate); propodus subequal in length to carpus; dactylus distinctly shorter than propodus; exopod flagellar, reaching midlength of merus; epipod curved, marginally minutely denticulate; podobranch about three-fourths length of epipod, simple. Major cheliped ( Fig. 3 View FIGURE 3 E) with merus bearing 1 minute denticle distally on dorsal margin, ventral margin serrated with 11 minute denticles; palm with 5 minute denticles on distal half of dorsal margin, lateral surface smooth; cusps on opposable margins of fingers much weaker than those in lectotype. Minor cheliped ( Fig. 3 View FIGURE 3 F) with ischium bearing 2 small denticles; merus with 3 tiny denticles on ventral margin; fixed finger with less acute and less conspicuous teeth on opposable margin. Uropodal exopod with lateral margin serrated with about 20 minute teeth; endopod with lateral margin serrated with 12 tiny acute teeth.
Myojin Knoll material. Generally similar to lectotype ( Figs 4 View FIGURE 4 , 5 View FIGURE 5 ). Rostrum with 3–4 low teeth on lateral margin (these teeth sharply pointed in 2 small specimens) ( Fig. 4 View FIGURE 4 C, D). Sixth abdominal somite with 4 denticles on posterodorsal margin ( Fig. 4 View FIGURE 4 G). Telson with 5–7 small teeth on lateral margin ( Fig. 4 View FIGURE 4 G). Third maxilliped as figured; ischium with crista dentata consisting of row of minute, sharply pointed denticles ( Fig. 5 View FIGURE 5 B). Major cheliped ( Fig. 5 View FIGURE 5 C) with ischium bearing 0–1 denticle on dorsal margin and 0–2 denticles on ventral margin; merus with 1 or 2 minute denticles on dorsal margin, ventral margin with 0–2 minute denticles; palm with 3–5 denticles on dorsal margin (in small specimen, these denticles acute, conspicuous). Minor cheliped ( Fig. 5 View FIGURE 5 D) with merus bearing 2 dorsal and 3 ventral denticles; palm with 1 minute denticle on dorsal margin; fixed finger with row of acute to subacute teeth on opposable margin. Male second pleopod with appendix masculina slightly shorter than appendix interna, bearing several stiff setae ( Fig. 4 View FIGURE 4 F). Uropodal exopod with lateral margin serrated with 11–14 tiny acute teeth; endopod with lateral margin serrated with 6–8 teeth.
Variation. As is apparent from the above description, the armature of the ventral margin of the merus of the major cheliped is variable, although this variation seems to be size-related. The numbers of denticles are as follows: 11 in the largest, female paralectotype (cl 6.9 mm), seven in the male lectotype (cl 5.0 mm), two in the non-type male from the Myojin Knoll (JAMSTEC 083387; cl 4.4 mm), and one in the male from the Myojin Knoll (JAMSTEC 83398; cl 3.9 mm). The third specimen from the Myojin Knoll (JAMSTEC 83713; cl 3.6 mm) has both chelipeds removed for DNA extraction by the second author, and thus the count of the denticles could not be recorded. The number of serrations on the lateral margin of the uropodal exopod shows similar pattern of sizerelated variation (six to up to 20).
Furthermore, the palm of the major cheliped is provided with low, tiny tubercles on the lateral and mesial surfaces in the male lectotype, whereas the surfaces of the palm are nearly smooth in the other specimens, including the female paralectotype. The rostrum is relatively broader in the female paralectotype than in the male specimens; and teeth on the opposable margin of the fixed finger of the minor cheliped are more acute in the male specimens than in the female paralectotype. It is still unclear whether these differences are attributable to sexual dimorphism.
Coloration in life. Body and appendages entirely whitish translucent. Eyestalk orange at base of cornea; cornea opaque.
Distribution and habitat. Banda Sea, Indonesia, 658 m ( Bate 1888), and Myojin Knoll, Izu Islands, Japan, 1074 m (this study). The newly collected specimens represent the second record of this species, as well as a new record for Japanese waters.
Ecology. The newly collected specimens were found to be associated with farreid sponge (Hexactinellida) collected at the inside slope of the Myojin Knoll Caldera located about 400 m from hydrothermal vent fields ( Fig. 6 View FIGURE 6 A). Two colonies of the sponge were collected, of which one and two shrimps (both males) were found inside the internal cavity respectively.
Remarks. Species of Eiconaxius Bate, 1888 are known to be associated with deep-water hexactinellid sponges (e.g., Ortmann 1891; Faxon 1893, 1896; Bouvier 1925; de Man 1925; Kensley 1996; Komai 2011). Valid species and their geographical ranges are summarized in Table 1. The genus is widely distributed in the Indo-Pacific, Gulf of Mexico and the Caribbean Sea, but the known range of each species is restricted to a rather narrow area. Species of the genus are morphologically similar ( Kensley 1996). Differences between species are generally minor, and assessment of variation is sometimes difficult because of the paucity of material. In fact, most species are still represented only by type material and few by subsequently reported specimens (see Sakai 2011). Kensley (1996) summarized characters of possible diagnostic significance.
The newly collected specimens from the Myojin Knoll, Japan, agree well with the type material except for the size-related variation in the number of ventral serrations on the merus of the major cheliped, as noted above.
The reexamination of the type material has revealed that Bate’s (1888) illustration is not altogether accurate. In particular, the rostrum of the male specimen (here designated as the lectotype) was figured as more elongate than in reality and with spinose lateral margins. Diagnostic characters of E. acutifrons , reassessed here, include: rostrum relatively narrow (1.4–1.9 times longer than wide), lanceolate; lateral margin of rostrum slightly to distinctly dentate; rostral median carina smooth, not bifurcate or hardly broadened posteriorly on gastric region of carapace; second and third abdominal pleura distinctly produced into an acute tooth, fourth pleuron also subacutely pointed ventrally; dactylus of major chela with cusp at midlength and fixed finger with opposing proximal cusp; and palm of major cheliped with minute denticles on dorsal margin, whereas that of minor cheliped unarmed or with one minute denticle. These characters are also shared by E. consobrinus and E. kermadeci compared below.
Eiconaxius consobrinus can be distinguished from E. acutifrons by the different structure of the chelipeds (de Man 1925). In E. consobrinus , the opposable margin of the fixed finger of the major cheliped has a deep notch just proximal to the prominent cusp arising at the midlength; the dactylar articulating margin of the palm of the minor cheliped is unarmed; and the dactylus of the minor cheliped is 1.1 times as long as the palm. On the other hand, in E. acutifrons , there is no prominent notch just proximal to the middle cusp on the opposable margin of the major cheliped; the dactylar articulating margin of the palm of the minor cheliped bears a bi-spined cusp on the lateral side; and the dactylus of the minor cheliped is about 1.5 times as long as the palm.
Eiconaxius kermadeci can be differentiated from E. acutifrons by the more distinctly delimited lateral rostral carinae, and the different configuration of the major chela. In E. kermadeci , there is a prominent hiatus between the fingers of the major chela, and the proximal part of the cutting edge of the fixed finger is armed with several strong teeth ( Bate 1888). As shown above, in E. acutifrons , there is no such a hiatus between fingers of the major chela and no strong proximal teeth on the cutting edge of the fixed finger.
Four species of Eiconaxius have been recorded the northwestern Pacific: E. farreae , E. kensleyi , E. mortenseni , and E. rubrirostris ( Ortmann 1891, Sakai 1992, Komai et al. 2010, Komai 2011). Of these, E. farreae is similar to E. acutifrons in the median carina on the carapace hardly bifurcate posteriorly and the relatively narrow, lanceolate rostrum. Nevertheless, E. farreae differs from E. acutifrons in the less produced and less acuminate posteroventral angles of the second and third abdominal pleura and the usually unarmed dorsal margin of the palm of the major cheliped. The lateral margins of the rostrum are more strongly upturned in E. farreae than in E. acutifrons .
Sakai (2011: 271) incorrectly cited the character of the male second pleopod in his diagnosis of E. acutifrons as “Male Plp 2 biramous, endopod with appendix interna, but without appendix masculina. [From Bate, 1888, pl. 5, fig. 2q].” According to Bate (1888: captions to the plate 5), the illustrated pleopod was the third pleopod, normally lacking an appendix masculina. In the descriptive text of the pleopods in the account of E. acutifrons, Bate (1888: 42) stated that “In the male these branches are smaller and less important, and the inner, instead of carrying one, supports two stylamblides.” This statement clearly indicates that the male second pleopod bore two appendices, i.e., appendices interna and masculina. No second pleopods of the male lectotype are now preserved, but the three additional male specimens all carry appendix masculina on the second pleopod.
Eiconaxius indonesicus View in CoL was established by Sakai (2011) for specimens previously referred to E. acutifrons View in CoL from Indonesia (De Man 1925), although he did not actually examine the types of E. acutifrons View in CoL or De Man’s (1925) specimens. Sakai (2011) cited the shape of the rostrum and armature of the major chela in differentiating E. indonesicus View in CoL from E. acutifrons View in CoL . Comparison between the present material of E. acutifrons View in CoL and the extensive description of de Man (1925) reveals the following morphological differences: the rostrum is broader in de Man’s (1925) specimens than in the types and additional specimens of E. acutifrons View in CoL ; the rostral apex forms a distinct point in de Man’s (1925) specimens, whereas there is no distinct point in the types and additional specimens of E. acutifrons View in CoL ; and the concavity distal to the proximal cusp on the opposable margin of the dactylus of the major cheliped is more prominent in de Man’s (1925) specimen than in the type specimens and additional specimens of E. acutifrons View in CoL . On the other hand, no significant difference in the spination of the dorsal margin of the major cheliped is seen between the type specimens of E. acutifrons View in CoL and the specimens of de Man (1925). Because of the abovementioned differences, we suppose at present that the taxon E. indonesicus View in CoL might be a valid species.
Kensley (1996) described a new species, E. albatrossae View in CoL , on the basis of specimens from Pacific Panama, referred by Faxon (1895) to E. acutifrons View in CoL . The type series consists of the following specimens: a single male from the Albatross station 3359 is the holotype; two males and four ovigerous females from the Albatross station 3358, and three males and two ovigerous females from Albatross station 3359 are paratypes ( Kensley 1996: 475). Sakai (2011) proposed a new taxon E. faxoni View in CoL for the same series of specimens, though he did not explicitly fix holotype or syntype (s). Consequently, Sakai’s taxon name is unavailable according to Article 16.4 and 72.3 of the ICZN (1999). Eiconaxius albatrossae View in CoL is also similar to E. acutifrons View in CoL , but the two species can be differentiated by the following characters ( Kensley 1996): the rostrum is relatively broader in E. albatrossae View in CoL than in E. acutifrons View in CoL ; the median gastric carina is distinctly bifurcated posteriorly in E. albatrossae View in CoL , but not bifurcated in E. acutifrons View in CoL ; the pleura of the second to fourth abdominal pleura are more strongly produced in E. albatrossae View in CoL than in E. acutifrons View in CoL ; the lateral margin of the telson bears more numerous teeth in E. albatrossae View in CoL than in E. acutifrons View in CoL (11 versus five to seven); and denticles on the dorsal margin of the palm of the major cheliped are more conspicuous in E. albatrossae View in CoL than in E. acutifrons View in CoL .
During this study, we have also reexamined two specimens from California (deposited in the LACM, one male and one female, 32°25’N, 119·60’W to 32°050’N, 119°65’E, 1200 m, 6 September 1978, coll. P. Gregory), identified as E. acutifrons View in CoL by Wicksten (1982), and found that they actually represent E. baja Kensley, 1996 View in CoL . Eiconaxius baja View in CoL is immediately recognized by the strongly upturned rostrum and the presence of strong setose tubercles on the lateral surface of the palms of chelipeds ( Kensley 1996).
Balss, H. (1925) Macrura der Deutchen Tiefsee-Expedition. 1. Palinura, Astacura und Thalassinidea. Wissenschaftliche Ergebnisse der Deutchen Tiefsee-Expedition auf dem Dampfer Valdivia , 1908 - 1910, 20, 189 - 216, pls 19, 18.
Bate, C. S. (1888) Report on the Crustacea Macrura collected by the Challenger during the years 1873 - 76. Report on the Scientific Results of the Voyage of H. M. S. Challenger during the years 1873 - 76, Zoology, 24, i - xc, 1 - 942, pls 1 - 157.
Borradaile, L. A. (1903) On the classification of the Thalassinidea. Annals and Magazine of Natural History, (7) 12, 534 - 551 + addendum on p. 638.
Bouvier, E. L. (1925) Les Macroures marcheurs. Reports on the results of dredging under the supervision of Alexander Agassiz in the Gulf of Mexico (1877 - 78), in the Caribbean Sea (1878 - 79), and along the Atlantic coast of the United States (1880), by the U. S. Coast Survey steamer Blake . Memoirs of the Museum of Comparative Zoology, Harvard University, 47, 401 - 472, pls 1 - 11.
Faxon, W. (1893) Reports on the dredging operations off the west coast of Central America to the Galapagos, to the west coast of Mexico, and in the Gulf of California, in charge of Alexander Agassiz, carried on by the U. S. Fish Commission Steamer Albatross during 1891, lieut. Commander Z. L. Tanner, U. S. N., commanding, 6: Preliminary description of new species of Crustacea. Bulletin of the Museum of Comparative Zoology at Harvard College, 24, 149 - 220.
Faxon, W. (1895) The stalk-eyed Crustacea: Reports on an exploration off the west coasts of Mexico, Central and South America, and off the Galapagos Islands, in charge of Alexander Agassiz, by the U. S. Fish Commission Steamer Albatross during 1891, Lieut. - Commander Z. L. Tanner, commanding, XV. Memoirs of the Museum of Comparative Zoology at Harvard College, 18, 1 - 292, pls A - H, J, K, 1 - 38, 38 bis, 39 - 56, 1 chart.
Faxon, W. (1896) Reports on the results of dredging, under the supervision of Alexander Agassiz, in the Gulf of Mexico and the Caribbean Sea, and on the east coast of United States, 1877 - 1880, by the U. S. coast survey steamer Blake, Lieut-Commander C. D. Sigsbee, U. S. N., and Commander J. R. Bartlett, U. S. N., commanding, 37: Supplementary notes on the Crustacea. Bulletin of the Museum of Comparative Zoology at Harvard College, 30, 153 - 168.
Hendrickx, M. (1995) Checklist of lobster-like decapod crustaceans (Crustacea: Decapoda: Thalassinidea, Astacidea and Palinuridea) from the eastern tropical Pacific. Annales del Instituto de Biologia, Universidad Nacional Autonoma de Mexico, Series Zoology, 66, 151 - 163.
ICZN (1999) Interntaional Code of Zoological Nomenclature. Fourth Edition. International Commission of Zoological Nomenclature, 306 pp.
Kensley, B. (1996) New thalassinidean shrimp from the Pacific Ocean (Crustacea: Decapoda: Axiidae and Calocarididae). Bulletin of Marine Science, 59, 469 - 489.
Komai, T., Lin, F. - J. & Chan, T. - Y. (2010) Five new species of Axiidae (Crustacea: Decapoda: Axiidea) from deep-water off Taiwan, with description of a new genus. Zootaxa, 2352, 1 - 28.
Komai, T. (2011) Deep-sea shrimps and lobsters (Crustacea: Decapoda: Dendrobranchiata and Pleocyemata) from the Sagami Sea and Izu Islands, central Japan. Memoirs of the National Museum of Nature and Science, 47, 279 - 337.
Man, G. J. de (1925) The Decapoda of the Siboga Expedition. Part VI. The Axiidae collected by the Siboga-Expedition. Siboga Expeditie Monographie, 39 a 5, 1 - 127.
Ortmann, A. (1891) Die Decapoden-Krebse des Strassburger Museums, mit besonderer Berucksichtigung der von Herrn Dr. Doderlein bei Japan und bei den Liu-Kiu-Inseln gesammelten und zur Zeit im Strassburger Museum aufbewahrten Formen. III. Die Abtheilungen der Reptantia Baos: Homaridea, Loricata und Thalassinidea. Zoologische Jahrbucher, Abtheilung fur Systematik, Geographie und Biologie der Thiere, 6, 1 - 58, pl. 1.
Sakai, K. & de Saint Laurent, M. (1989) A check list of Axiidae (Decapoda, Crustacea, Thalassinidea, Anomura), with remarks and in addition descriptions of one new subfamily, eleven new genera and two new species. Naturalists, Publications of Tokushima Biological Laboratory, Shikoku University, 3, 1 - 104.
Sakai, K. (1992) Axiid collections of the Zoological Museum, Copenhagen, with the description of one new genus and six new species (Axiidae, Thalassinidea, Crustacea). Zoologica Scripta, 21, 157 - 180.
Sakai, K. (2011) Axioidea of the world and a reconsideration of the Callianassoidea (Decapoda, Thalassinidea, Callianassida). Crustaceana Monographs, 13, i - x, 1 - 616.
Wicksten, M. K. (1982) Crustaceans from baited traps and gill nets off southern California. California Fish and Game, 68, 244 - 248.
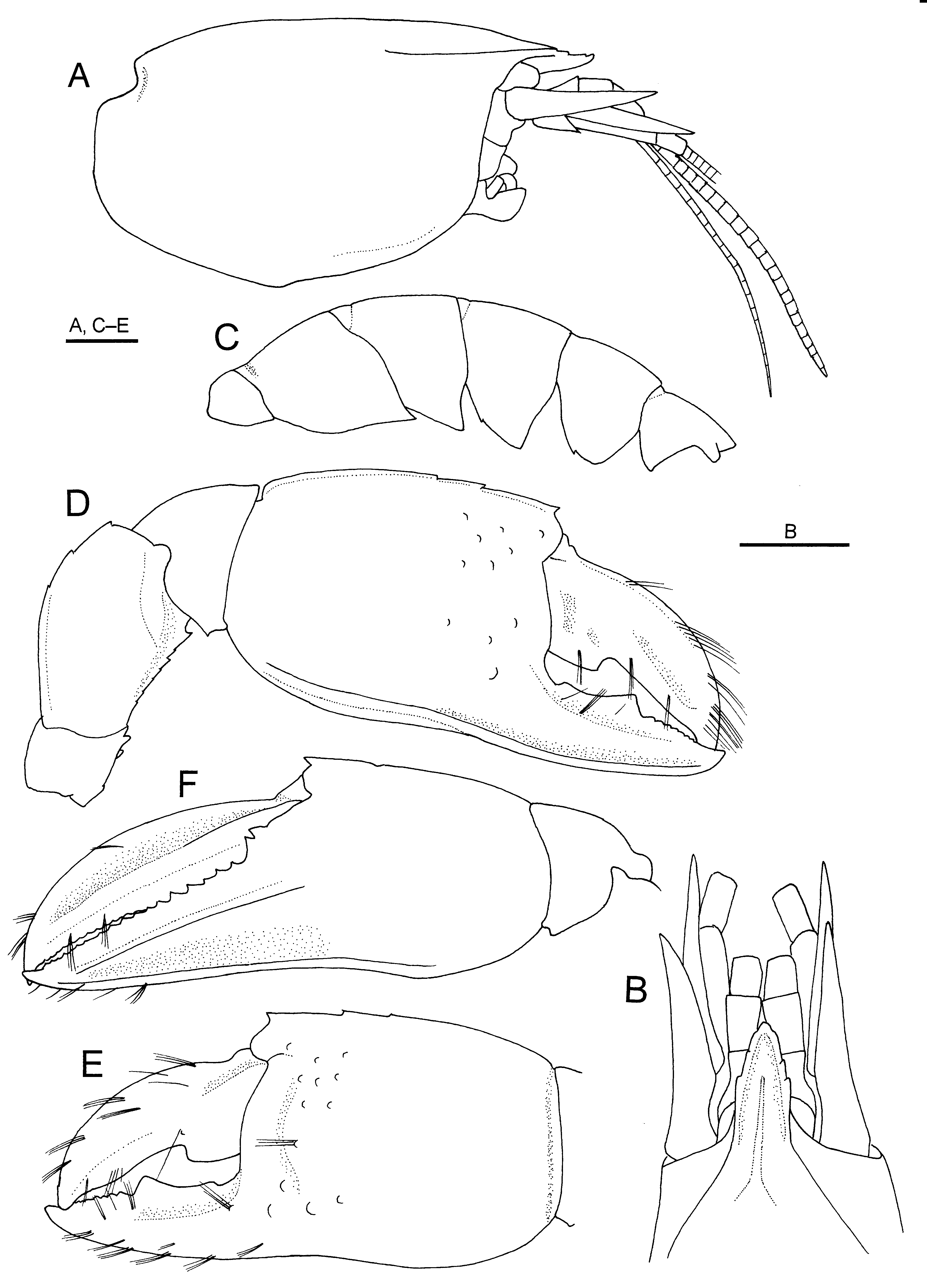
FIGURE 1. Eiconaxius acutifrons Bate, 1888, lectotype, male (cl 5.0 mm), NHM 1888: 22. A, carapace and cephalic appendages, right side, lateral view; B, anterior part of carapace and cephalic appendages, dorsal view (antennular and antennal flagella omitted); C, abdomen, left side, lateral view; D, right major cheliped, lateral view; E, same, chela, mesial view; F, chela and carpus of left minor cheliped, lateral view. Scale bar: 1 mm.
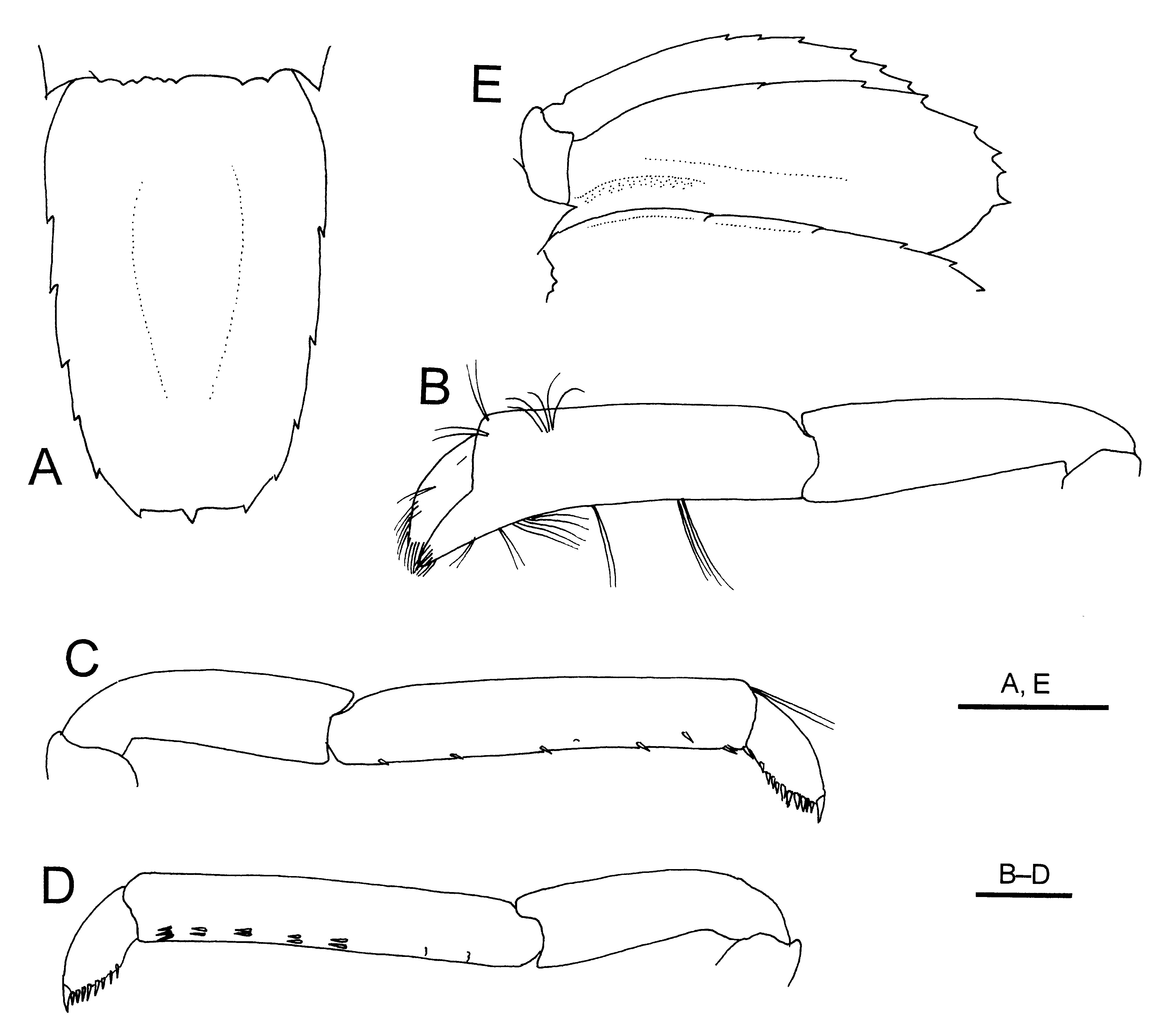
FIGURE 2. Eiconaxius acutifrons Bate, 1888, lectotype, male (cl 5.0 mm), NHM 1888: 22. A, telson, dorsal view; B, chela and carpus of left second pereopod, lateral view; C, carpus to dactylus of right third pereopod, lateral view; D, carpus to dactylus of left fourth pereopods, lateral view. Scale bars: 1 mm for A, E; 0.5 mm for B – D.
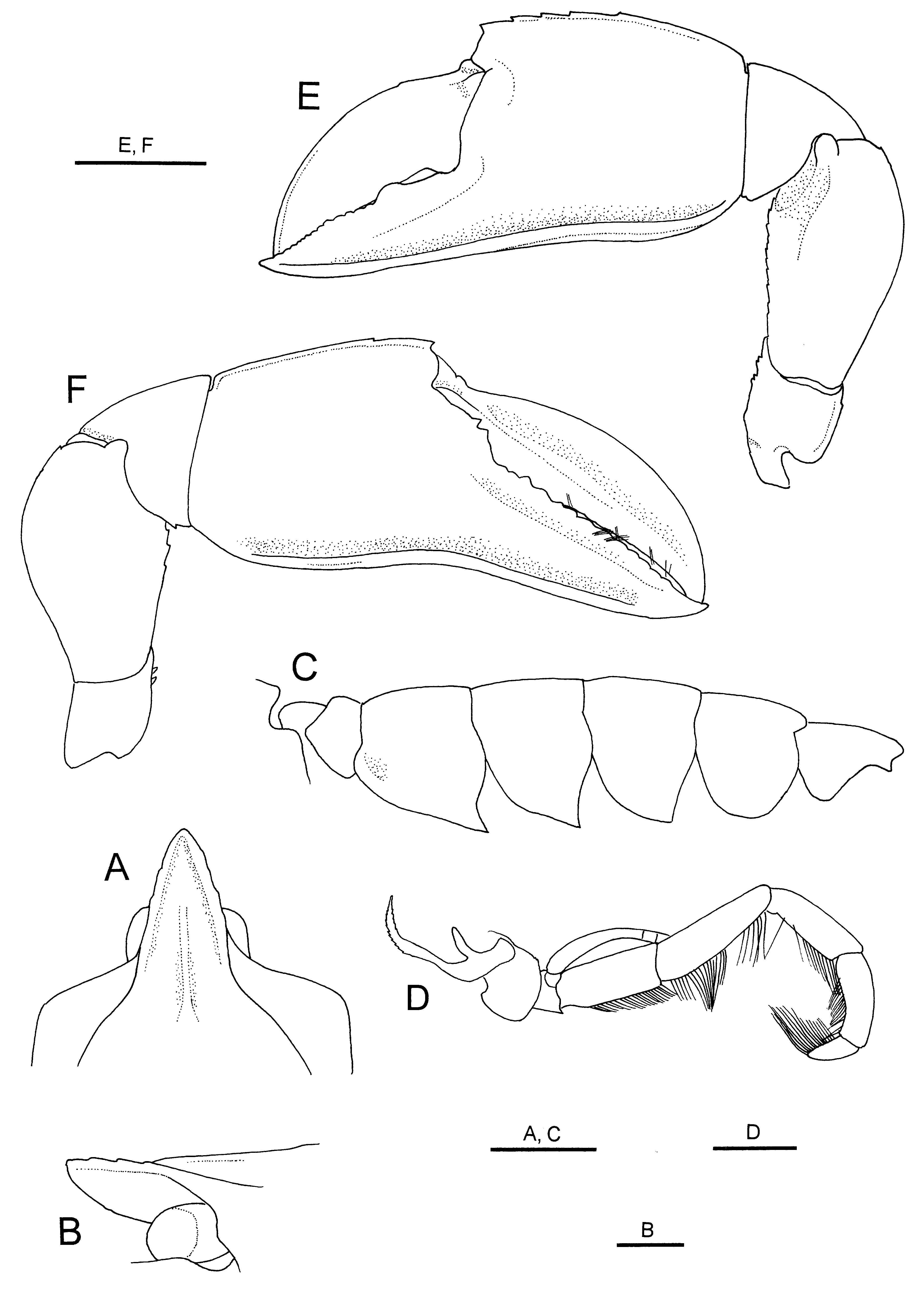
FIGURE 3. Eiconaxius acutifrons Bate, 1888, paralectotype, ovigerous female (cl 6.9 mm), NHM 1888: 22. A, anterior part of carapace and eyes, dorsal view; B, same, lateral view; C, right third maxilliped, lateral view; E, left major cheliped, lateral view; F, right minor cheliped, lateral view. Scale bars: 2 mm for C, E, F; 1 mm for A, D; 0.5 mm for B.
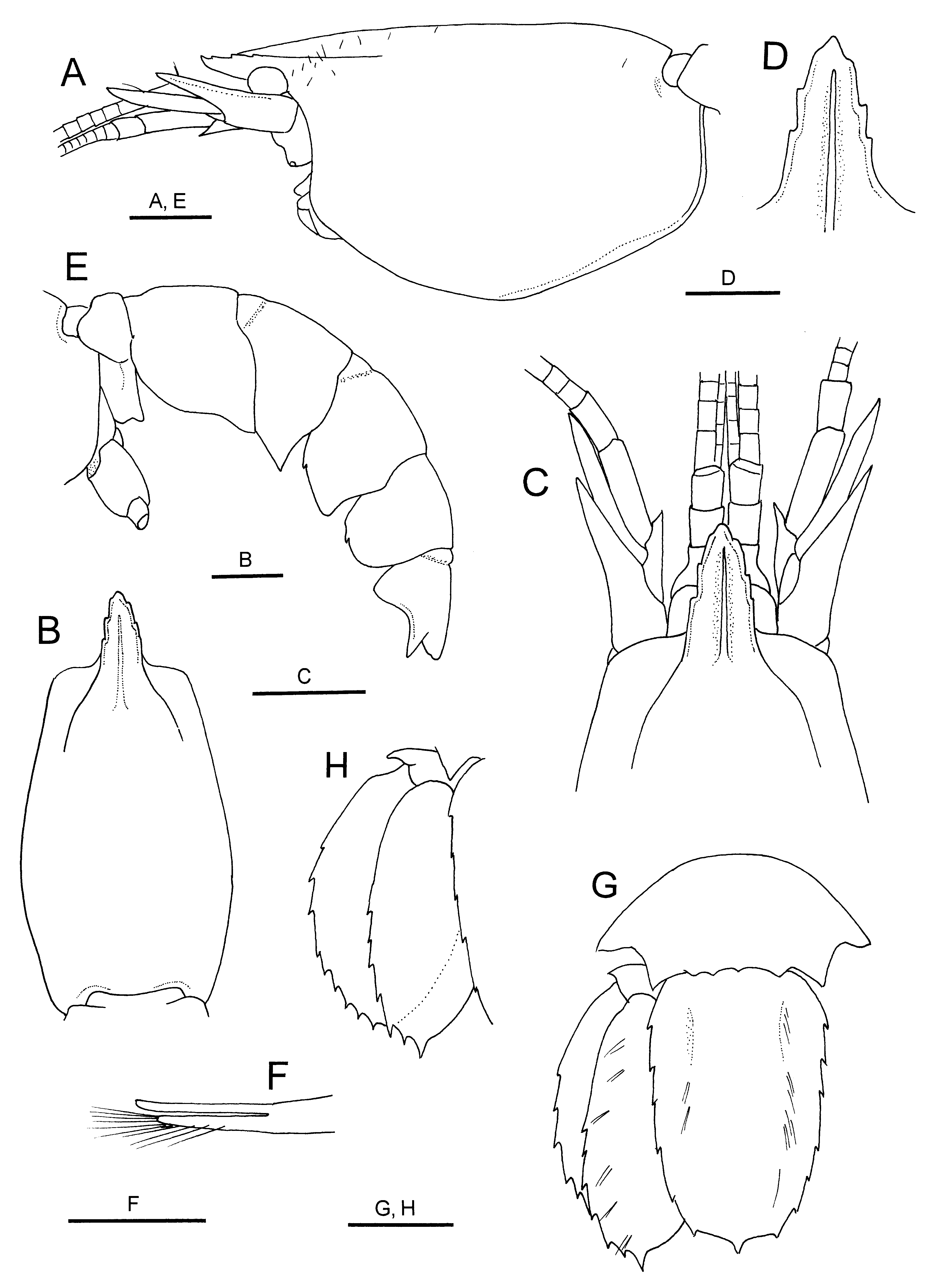
FIGURE 4. Eiconaxius acutifrons Bate, 1888, non-type, male (cl 4.5 mm), JAMSTEC 0 83387. A, carapace and cephalic appendages, lateral view; B, carapace, dorsal view; C, anterior part of carapace and cephalic appendages, dorsal view; D, rostrum, dorsal view; E, abdomen, lateral view; F, appendices interna and masculina of left second pleopod, dorsolateral view; G, sixth abdominal somite, telson and left uropod, dorsal view; H, left uropod, dorsal (perpendicular) view. Scale bars: 1 mm for A – C, E, G, H; 0.5 mm for D, F.
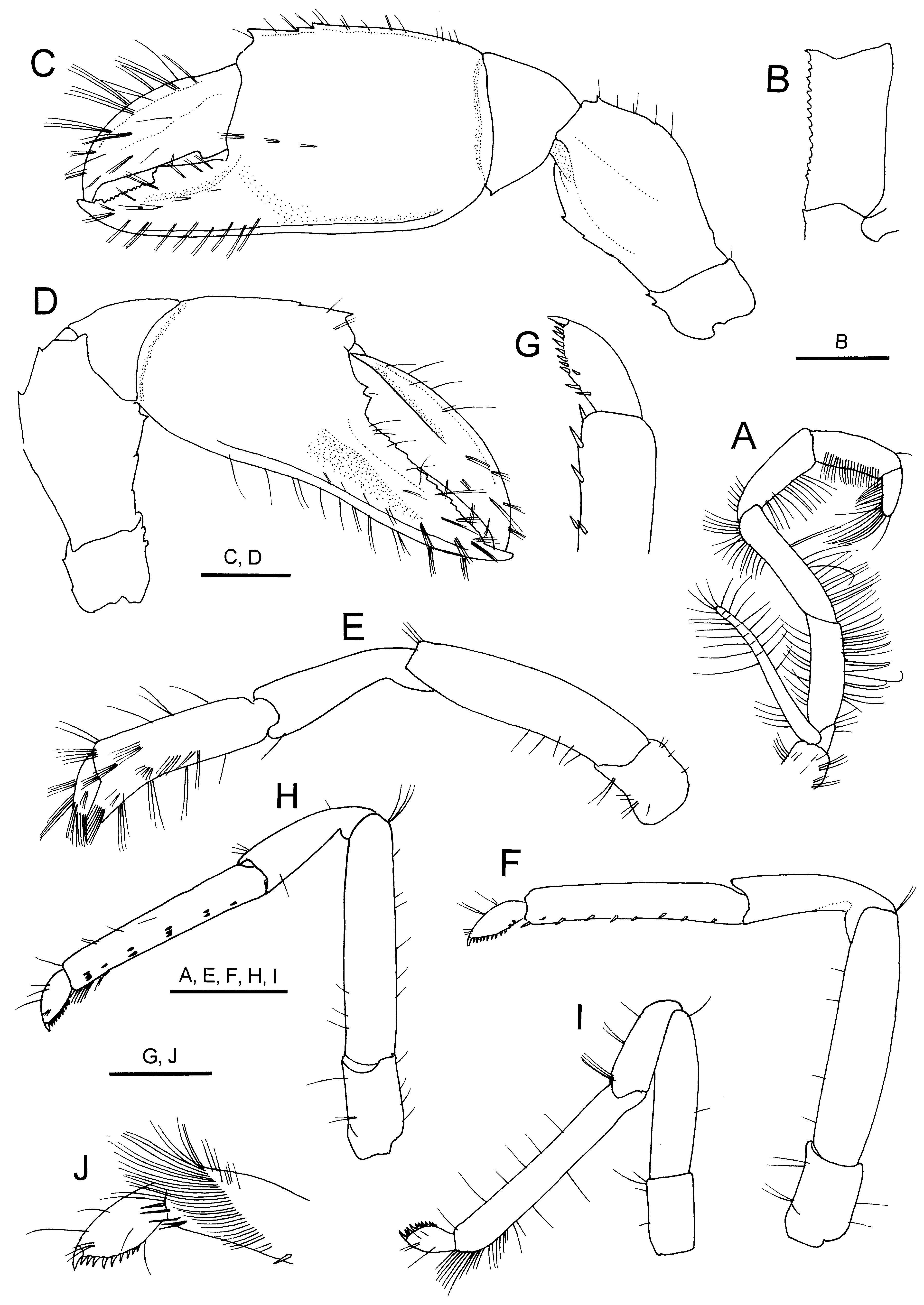
FIGURE 5. Eiconaxius acutifrons Bate, 1888, non-type, male (cl 4.5 mm), JAMSTEC 0 83387. A, right third maxilliped; B, same, ischium, dorsal view; C, left major cheliped, lateral view; D, right minor cheliped; E, left second pereopod, lateral view; F, left third pereopod, lateral view; G, same, distal part of propodus and dactylus; H, left fourth pereopod, lateral view; I, left fifth pereopod, lateral view; J, same, distal part of propodus and dactylus. Scale bars: 1 mm for A, C – F, H, I; 0.5 mm for B, G. J.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eiconaxius acutifrons Bate, 1888
| Komai, Tomoyuki & Tsuchida, Shinji 2012 |
Axius acutifrons
| Hendrickx 1995: 157 |
| Wicksten 1982: 246 |
Iconaxius acutifrons
| Balss 1925: 209 |
Axius (Eiconaxius) acutifrons
| Man 1925: 37 |
Axius (Eiconaxius) acutifrons:
| Borradaile 1903: 538 |
Axius acutifrons
| Faxon 1895: 103 |
Eiconaxius acutifrons
| Sakai 2011: 270 |
| Kensley 1996: 475 |
| Saint 1989: 16 |
| Bate 1888: 40 |
1 (by plazi, 2016-04-12 22:29:05)
2 (by ImsDioSync, 2016-12-21 04:19:39)
3 (by ImsDioSync, 2016-12-21 04:20:20)
4 (by ImsDioSync, 2017-06-22 02:34:55)
5 (by ImsDioSync, 2017-06-22 03:02:45)
6 (by ExternalLinkService, 2019-09-26 19:21:55)
7 (by ExternalLinkService, 2022-01-30 13:22:11)
8 (by ExternalLinkService, 2022-02-20 08:33:37)
9 (by plazi, 2023-10-26 09:12:45)