
Tetramorium striativentre Mayr, 1877
|
publication ID |
https://doi.org/ 10.1515/vzoo-2015-0024 |
|
DOI |
https://doi.org/10.5281/zenodo.6461989 |
|
persistent identifier |
https://treatment.plazi.org/id/03AAA223-B07B-6911-42BE-78B7076EFA22 |
|
treatment provided by |
Felipe |
|
scientific name |
Tetramorium striativentre Mayr, 1877 |
| status |
|
Tetramorium striativentre Mayr, 1877 View in CoL
Tetramorium caespitum var. striativentre Mayr, 1877: 16 View in CoL , worker, ♀ (part., worker from Samarkand and ♀), Uzbekistan (redescribed in German: Mayr, 1880: 35) (see also Taxonomic notes, below). Tetramorium caespitum subsp. striativentre: Ruzsky, 1905 a: 538 View in CoL . Tetramorium striativentre: Dalla Torre, 1893: 135 View in CoL ; Ruzsky, 1905 a: 766; 1905 b: 518; Emery, 1909: 706; Ruzsky, 1923: 4; Collingwood, 1961 a: 54; 1961 b: 289; Pisarski, 1967: 403; Dlussky, Zabelin, 1985: 232; Dlussky et al., 1990: 208; Radchenko, 1992 a: 45; 1992 b: 52; Collingwood, Heatwole, 2000: 10; Paknia et al., 2010: 35.
M a t e r i a l e x a m i n e d. Lectotype, worker (designated here), “Samarkand” (original label of A. Fedchenko is in Russian — “Самаркандъ”), “7” ( ZMMU) . Non-type material. Ca. 100 workers, 7 ♀, 2 Ơ from Uzbekistan, Turkmenistan, Kyrgyzstan, Tajikistan, Afghanistan and Iran .
Measurements of workers (in mm; n = 30), ordered as: lectotype (min-max) [mean ± SD] (see also fig. 6 View Fig , a–d): HL 0.77 (0.77–0.91) [0.86 ± 0.041], HW 0.76 (0.73–0.85) [0.81 ± 0.037], FW 0.28 (0.28–0.35) [0.32 ± 0.007], FLW 0.29 (0.29–0.35) [0.32 ± 0.014], OL 0.17 (0.15–0.21) [0.18 ± 0.013], GnL 0.18 (0.18–0.25) [0.23 ± 0.015], SL 0.62 (0.55–0.64) [0.60 ± 0.025], ML 0.98 (0.92–1.15) [1.07 ± 0.072], PNW 0.56 (0.48–0.59) [0.54 ± 0.032], PL 0.34 (0.31–0.36) [0.33 ± 0.016], PW 0.22 (0.22–0.32) [0.29 ± 0.026], PH 0.27 (0.25–0.32) [0.29 ± 0.020], PndL 0.18 (0.18–0.24) [0.21 ± 0.019], PPL 0.22 (0.19–0.27) [0.22 ± 0.013], PPW 0.27 (0.27–0.35) [0.31 ± 0.020], PPH 0.25 (0.23–0.32) [0.28 ± 0.016], ESL 0.14 (0.04–0.14) [0.08 ± 0.028], ESD 0.12 (0.12–0.25) [0.21 ± 0.027], HTL 0.56 (0.55–0.64) [0.61 ± 0.015], rug-frons 21 (16–21) [18 ± 0.252].
Рис. 6. Детали строения Tetramorium striativentre , рабочий: a — тело в профиль; b — грудь и стебелек, вид сверху; c — голова, вид сверху; d — брюшко, вид сверху.
Indices: CI 1.01 (1.01–1.11) [1.07 ± 0.026], SI 1 0.81 (0.67–0.81) [0.70 ± 0.029], SI 2 0.82 (0.71–0.82) [0.75 ± 0.027], FI 0.37(0.37–0.42) [0.40 ± 0.014], FLI 1.04 (1.00–1.06) [1.01 ± 0.016], OI 1 0.22 (0.20–0.26) [0.23 ± 0.011], OI 2 0.94 (0.68–0.94) [0.81 ± 0.055], PI 1 1.26 (1.03–1.27) [1.16 ± 0.060], PI 2 0.29 (0.29–0.39) [0.36 ± 0.020], PndI 1.22 (1.21–1.48) [1.34 ± 0.063], PPI 1 0.81 (0.60–0.90) [0.72 ± 0.038], PPI 2 0.36 (0.35–0.42) [0.38 ± 0.019], ESLI 0.18 (0.05–0.18) [0.10 ± 0.034].
Measurements of queens (in mm; n = 7), ordered as: min-max [mean ± SD] (see also fig. 7 View Fig , a–c): HL 1.05–1.14 [1.10 ± 0.033], HW 1.05–1.14 [1.09 ± 0.032], FW 0.40–0.48 [0.44 ± 0.032], FLW 0.39–0.47 [0.42 ± 0.033], OL 0.29–0.33 [0.30 ± 0.012], GnL 0.22–0.26 [0.24 ± 0.014], SL 0.72–0.78 [0.74 ± 0.022], PL 0.46–0.51 [0.48 ± 0.016], PW 0.43–0.51 [0.46 ± 0.023], PH 0.43–0.48 [0.46 ± 0.023], PndL 0.23–0.27 [0.25 ± 0.018], PPL 0.26–0.32 [0.29 ± 0.021], PPW 0.49–0.56 [0.53 ± 0.024], PPH 0.43–0.49 [0.46 ± 0.027], ESL 0.05–0.13 [0.08 ± 0.027], ESD 0.39–0.43 [0.42 ± 0.016], HTL 0.73–0.81 [0.77 ± 0.026], rug-frons 18–
Рис. 7. Детали строения Tetramorium striativentre , самка; a — тело в профиль; b — тело, вид сверху; c — голова, вид сверху.
23 [21 ± 1.902], ML 1.73–1.93 [1.80 ± 0.081], MH 0.90–0.98 [0.93 ± 0.030], SCL 1.24–1.36 [1.29 ± 0.040], SCW 0.96–1.04 [0.98 ± 0.027].
Indices: CI 0.97–1.04 [1.01 ± 0.023], SI 1 0.66–0.68 [0.67 ± 0.009], SI 2 0.64–0.68 [0.67 ± 0.020], FI 0.36–0.46 [0.41 ± 0.030], FLI 0.91–0.97 [0.96 ± 0.025], OI 1 0.26–0.28 [0.27 ± 0.008], OI 2 1.21–1.33 [1.26 ± 0.045], PI 1 0.95–1.12 [1.04 ± 0.062], PI 2 0.38–0.48 [0.42 ± 0.032], PndI 1.67–1.90 [1.78 ± 0.094], PPI 1 0.50–0.61 [0.54 ± 0.042], PPI 2 0.46–0.53 [0.49 ± 0.021], ESLI 0.05–0.12 [0.08 ± 0.025], MI 1.90–2.03 [1.95 ± 0.043] SCI 1.28–1.35 [1.31 ± 0.028].
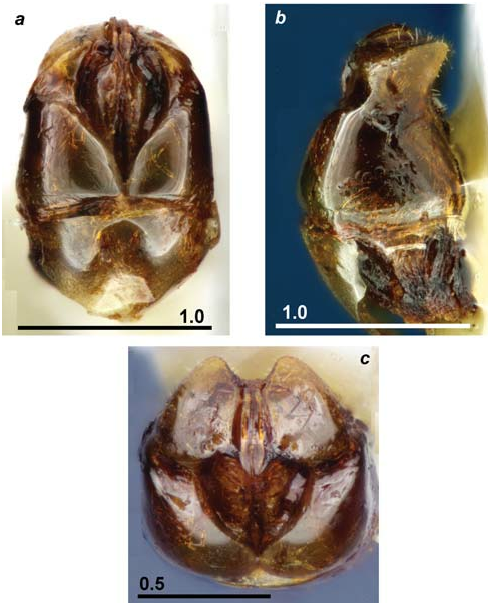
First description of males ( fig. 8 View Fig , a–c; 9 View Fig , a–c).
M a t e r i a l e x a m i n e d. 2 Ơ, Uzbekistan, Aman-Kurgan , 25.05.1942, leg. K. Arnoldi ( ZMMU) .
Head somewhat wider than length, broadly rounded above eyes, occipital margin convex. Anterior clypeal margin gradually convex, not prominent. Eyes big, situated distinctly below midlength of sides of head, so that genae quite short. Scape length about half of head width. Mandibles with five sharp teeth. Sculpture on head dorsum not very coarse (distinctly less coarse than in T. sabatinellii ): frons with longitudinal and longitudinallyconcentric rugosity, clypeus with semi-concentric rugosity, surface between and behind lateral ocelli with not coarse transversal rugae, remaining part of head dorsum quite coarsely and densely punctated. Occipital margin and temples with not abundant long erect hairs.
Mesosoma long and low, scutum and scutellum convex, notauli well developed. Petiole not very long and quite low, with almost straight anterior face and rounded node dorsum. Petiole widened, its node wider than length (seen from above), postpetiole <1.4 times wider than petiole. Scutum and scutellum with dense longitudinal rugulosity, surface between rugulae densely but not coarsely punctated, while lateral parts of dorsum of scutum smooth and shiny. Mesopleura and propodeum densely, but not coarsely longitudinally rugulose, pronotum mostly smooth. Petiolar node and postpetiole densely and coarsely punctated, dorsum of petiolar node also with longitudinally-concentric rugae.
Stipites of genitalia gradually curved inward apically (seen dorsally or ventrally), their apices slightly excavated (seen from behind).
Mesosoma and waist with not abundant long erect hairs. Surface of gastral tergites completely smooth and shiny. Body colour blackish-brown, mandibles and appendages brownish-red.
Measurements of males (in mm; n = 2), ordered as: min-max: HL 0.70–0.72, HW 0.73–0.73, OL 0.31–0.33, GnL 0.08–0.08, SL 0.36–0.38, PL 0.36–0.36, PW 0.31–0.33, PH 0.29–0.30, PPL 0.26–0.26, PPW 0.43–0.44, PPH 0.36–0.39, HTL 0.95–0.96, ML 1.91–1.98, MH 1.18–1.24, SCL 1.40–1.40, SCW 0.95–0.98.
Indices: CI 0.96–0.98, SI 1 0.51–0.54, SI 2 0.50–0.52, OI 1 0.43–0.45, OI 2 4.00–4.17, PI 1 1.22–1.27, PI 2 0.43–0.45, PPI 1 0.59–0.61, PPI 2 0.59–0.61, MI 1.55–1.67, SCI 1.44–1.48.
T a x o n o m i c n o t e s. T. striativentre was described by Mayr (1877, in Russian; redescribed in German by him in 1880) based on 2 workers and 1 queen, collected by the expedition of A. P. Fedchenko. The main diagnostic feature of this species is the sculpture of the first gastral tergite that is completely densely longitudinally striato-punctated. As the type localities Mayr (1877: 17) recorded: “7th of March 1869 in Samarkand, 9th of May in Ulus, beginning of May 1871 in Kisil-Kumy“ (now Uzbekistan). Unfortunately, it is not obvious from the original description, where from workers and queen are originated. Moreover, Mayr (loc. cit., p. 16) noted, that the propodeum of one worker specimen is with “two short, barely visible denticles, while the second worker has spines, length of which is distinctly longer than their width at the base” (our translation from Russian).
Ruzsky (1905 a: 766) investigated the syntypes worker and queen of T. striativentre preserved at that time in the collection of ZMMU, and confirmed heterospecificity of this species and T. schneideri . Unfortunately, he also did not provide any label data for these specimens, but noted that worker specimen has original Mayr’s label. Nowadays we found in the collection of ZMMU only one worker from the Mayr’s type series with the original labels from the collection of Fedchenko: “Самаркандъ”, “7”. We designated this worker as the lectotype of T. striativentre , despite it seems somewhat “atypical” by some features compare to all other examined specimens of this species from its whole range (about 100 workers). Particularly, it has much longer propodeal spines and relatively longer scape what is similar to those of T. schneideri . Nevertheless, by the most important diagnostic feature, such as the shape of the frontal carinae and lobes, the character of sculpture of the head and waist, and the shape of the petiolar node it certainly belongs to T. striativentre (see below).
Dlussky et al. (1990) pointed out that type specimens (Sic!: perhaps it is misprinting, as it should be only one worker) of T. striativentre from Kisil-Kumy, preserved in the collection of ZMMU, belong to T. schneideri . Unfortunately, we could not find corresponding material in that collection (investigated in 2012). Most probably, it is a specimen with longer spines, mentioned by Mayr (see above).
Preceding authors (e. g. Ruzsky, 1905 a, b; Dlussky et al., 1990) had used for the separation of T. striativentre and T. schneideri first of all the character of sculpture on the head dorsum and the length of the propodeal teeth. These features are normally useful, but occasionally they may be quite variable and sometimes single specimen of both species may be hardly distinguishable. Thus, despite the sculpture on the head dorsum in T. striativentre is generally coarser than in T. schneideri , the number of longitudinal rugae level with the eyes in the first species is 16–21 (mean 18) and 17–33 (mean 23) it the latter one; at the same time, the length of propodeal spines are rather variable in both species, despite they are generally shorter in T. striativentre (mean ESLI 0.10 with the range 0.05–0.18 vs. mean 0.19 with the range 0.15–0.25 in T. schneideri ).
Radchenko (1992 a) added as an important feature for the separation of these species the shape of the petiolar node: transversal in T. striativentre (PndI> 1.20) and subcircle in T. schneideri (PndI <1.15).
In the course of the current revision we paid an attention to one more very distinct feature never used before for the separation of T. striativentre and all other species with the completely striato-punctated first gastral tergite (e. g. T. schneideri , T. saudicum and T. sabatinellii ): the frontal carinae in T. striativentre are not curved, gradually converging anteriorly so that the frontal lobes are not extended (FLI 1.00–1.06, mean 1.01), but in three other species the frontal carinae are curved above the antennal insertions and the frontal lobes are distinctly extended (FLI ≥ 1.08, means 1.12 … 1.17) (compare fig. 6 View Fig , c and 1 View Fig , c; 10 View Fig , c).
Finally, T. striativentre well differs from three above mentioned species by the more regular, longitudinally-concentric rugae on the dorsum of the petiolar node and postpetiole (compare fig. 6 View Fig , b and 1 View Fig , b; 10 View Fig , b).
D i s t r i b u t i o n. Central Asia, Afghanistan, Iran, NW China. André (1883) has recorded this species form Palestine (Nazareth), and Wheeler and Mann (1916) — from Syria and Jordan. We did not see the corresponding material, but based on the known distribution of T. striativentre we may suspect that records from Palestine, Syria and Jordan might concern T. sabatinellii .
E c o l o g y. It inhabits mostly mountain regions where prefers steppe-like and semidesert biotopes with sparse vegetations. Nests are built in a soil, often under stones. Zoonecrophagic species, collecting both living small invertebrates and their remnants, but quite actively collects also plant seeds. Nuptial flights occur in the end of May or in June (depending from region) (for more details see Dlussky, 1981, Dlussky et al., 1990).





| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tetramorium striativentre Mayr, 1877
| Radchenko, A. G. & Scupola, A. 2015 |
Tetramorium caespitum var. striativentre
| Paknia, O. & Radchenko, A. & Pfeiffer, M. 2010: 35 |
| Collingwood, C. A. & Heatwole, H. 2000: 10 |
| Radchenko, A. G. 1992: 45 |
| Radchenko, A. G. 1992: 52 |
| Dlussky, G. M. & Soyunov, O. S. & Zabelin, S. I. 1990: 208 |
| Dlussky, G. M. & Zabelin, S. I. 1985: 232 |
| Pisarski, B. 1967: 403 |
| Ruzsky, M. 1923: 4 |
| Emery, C. 1909: 706 |
| Ruzsky, M. 1905: 538 |
| Ruzsky, M. 1905: 766 |
| Ruzsky, M. 1905: 518 |
| Dalla Torre, C. G. 1893: 135 |
| Mayr, G. 1880: 35 |
| Mayr, G. 1877: 16 |