
Cotithene gorayebi Valente, da Silva & de Medeiros, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4576.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:7A829CA9-A8B3-4951-A124-2B316585AE8A |
|
DOI |
https://doi.org/10.5281/zenodo.3717699 |
|
persistent identifier |
https://treatment.plazi.org/id/03A987C9-4A67-FFD4-E1B1-FCFFF03CFE87 |
|
treatment provided by |
Plazi |
|
scientific name |
Cotithene gorayebi Valente, da Silva & de Medeiros |
| status |
sp. nov. |
Cotithene gorayebi Valente, da Silva & de Medeiros , sp. nov.
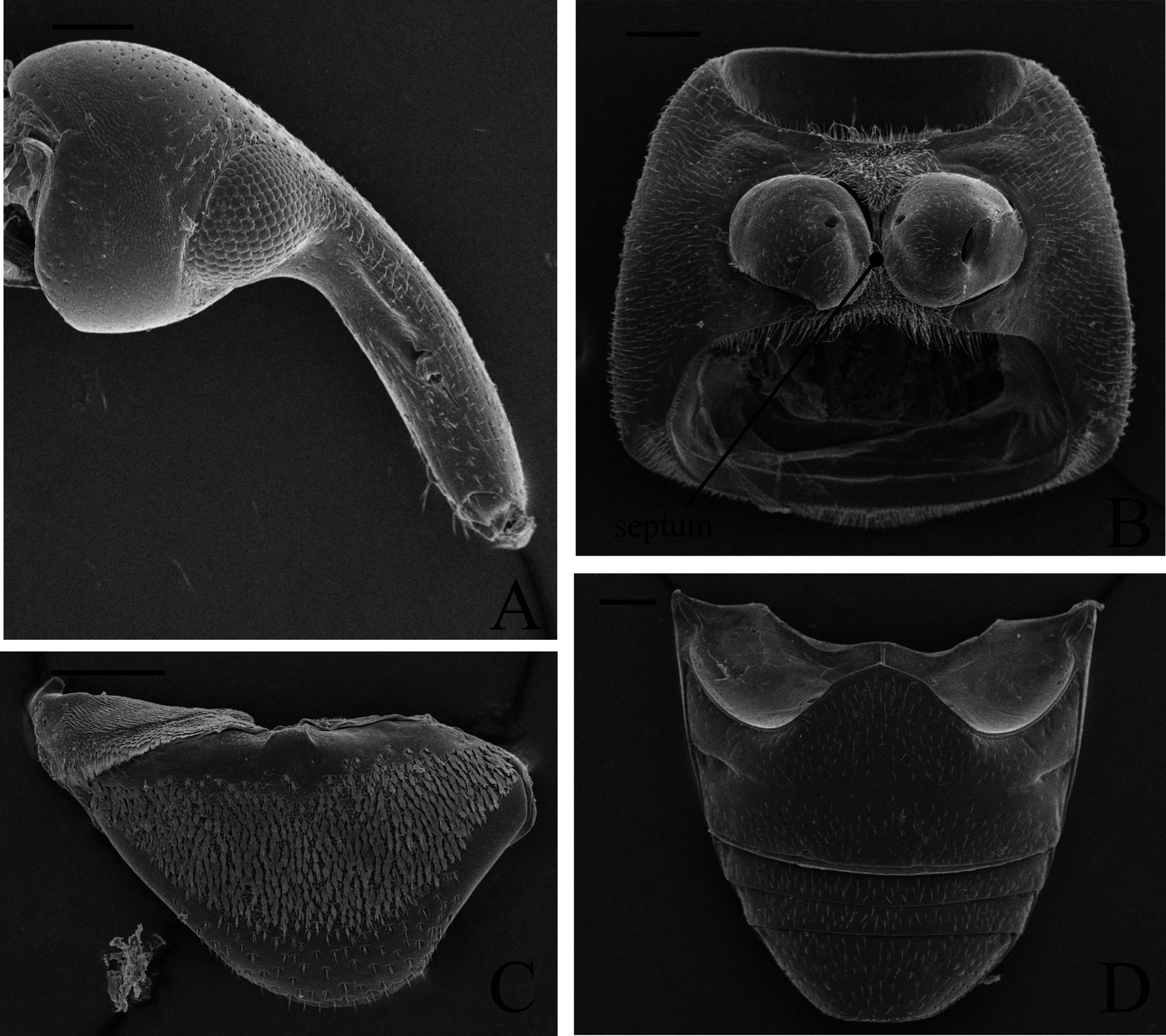
( Figs 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 , 9–12 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 )
Diagnosis. This species differs from other described species of the genus by its smaller size (male: 1.9–2.2 mm, female: 1.6–2.1 mm); integument color pattern dissimilar between sexes ( Fig. 2 View FIGURE 2 , see details below); procoxal cavities separated by only a narrow septum in both male and female ( Figs 2 View FIGURE 2 , 3D View FIGURE 3 , 6B View FIGURE 6 ); elytral stria I sulcate and; remaining striae very shallow and very similar to elytral intervals ( Figs 2 View FIGURE 2 , 4D View FIGURE 4 ). Males are also distinguished by the aedeagus ( Figs 5 View FIGURE 5 D–H) bearing two inner separately arched rami dorsally projected on each side (in the remaining species each side with only one separated arched ramus) and by the endophallus of the aedeagus with a basal sclerite. Females are also distinguished by the sternum VIII ( Fig. 7C View FIGURE 7 ) with a pentagonal-shaped lamina and furcal arms connected only by membranes, not closed and bearing only subequal setae on apices.
Similar species: Females of Cotithene dicranopygia Franz and Cotithene stratiotricha have procoxal cavities separated by a narrow septum as in the new species, but females of Cotithene gorayebi can be distinguished from both by the shape of sternum VIII, and also from C. stratiotricha by a C-shaped spermatheca. Males of Cotithene anaphalanta Franz and Cotithene melanoptera can superficially resemble males of the new species due the elytra evidently dark reddish brown to black but males of Cotithene gorayebi have the rostrum and head darker reddish brown to black in addition to dark elytra. Also, Cotithene gorayebi and C. melanoptera are associated with inflorescences of Evodianthus funifer (for details, see Natural History) but the new species has been recorded in Brazil (Pará) while C. melanoptera has been recorded only in Venezuela (Merida and Aragua).
Males of Cotithene can be characterized by the rostrum variously expanded near the apex to basal 2/5 (see Franz 2008). In Cotithene gorayebi ( Figs 2A View FIGURE 2 , 3 View FIGURE 3 A–B), as in Cotithene dicranopygia and Cotithene stratiotricha , the males have the rostrum angulate-tumescent near basal 2/5; however, the rostrum of the new species also bears a tubercle on the inner margin of the eye ( Figs 2A View FIGURE 2 , 3A View FIGURE 3 ) and a irregular dorsolateral carina ( Figs 3 View FIGURE 3 A–B). These sexually dimorphic features (tumescence, tubercles and carinae) on the male rostrum of Cotithene gorayebi are very conspicuous in the largest males but nearly indistinct in the smallest males, with intermediate states present as well. Furthermore, large males have a larger and more inflated pronotum. Franz (2008) observed similar variations in males of C. stratiotricha , Cotithene globulicollis Voss and C. melanoptera . He considered these scaled modifications as apparent positive allometry, possibly related to alternative reproductive strategies of the males for access to reproductively active females. Here we also observed males of Cotithene gorayebi making use of the rostrum and head in male-male conflicts during the approach of females. Thus the scaled modifications on the rostrum and pronotum observed in males of Cotithene gorayebi apparently represent a case of positive allometry as well.
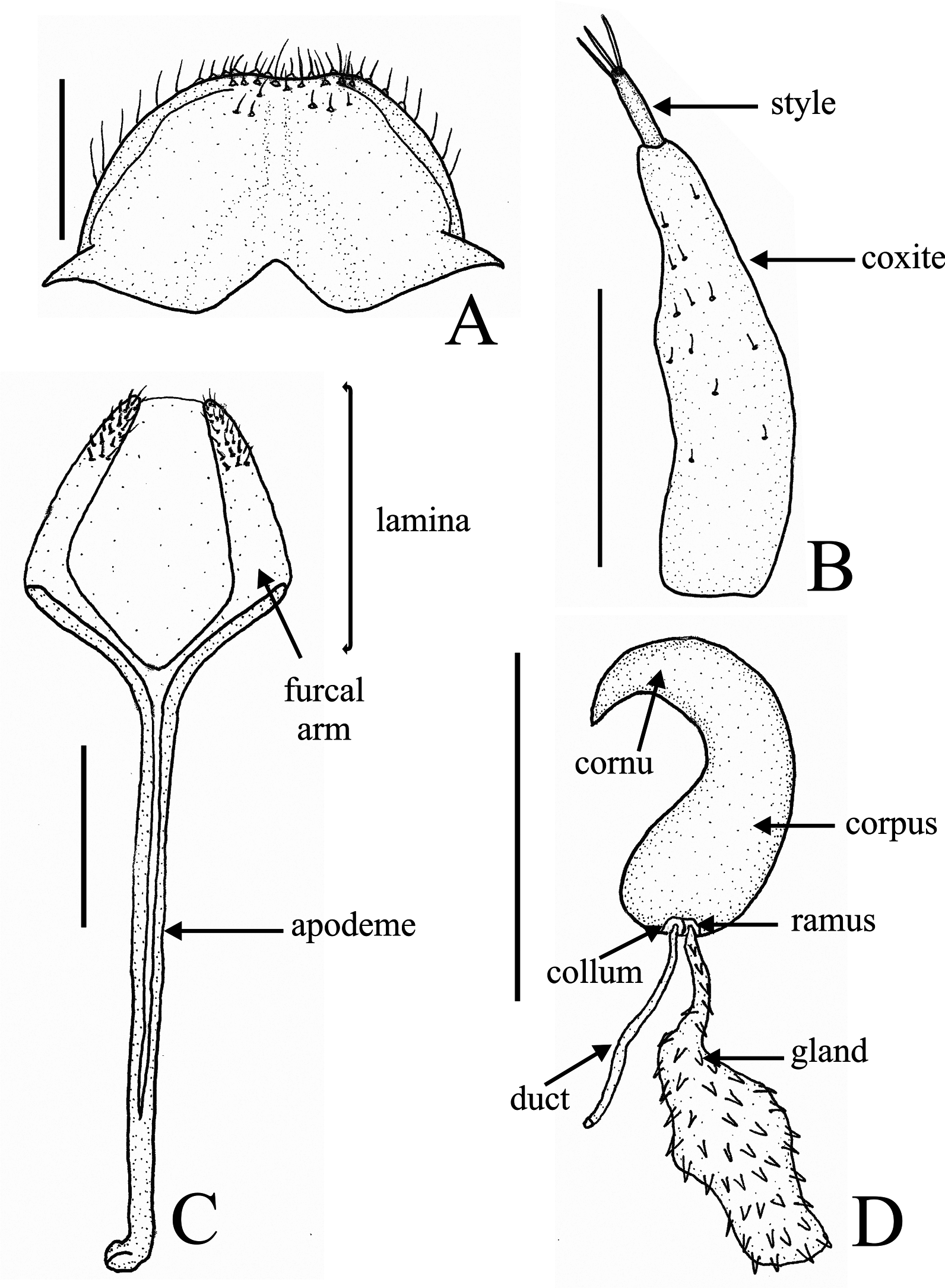
Description. Male ( Figs 2A View FIGURE 2 , 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). Length 1.9–2.2 mm, width 0.8–1.0 mm, elongated l/w = 2.3–2.5. Integument ( Fig. 2A View FIGURE 2 ) from light reddish brown to orange with head, rostrum, elytra (except margins) and anterior margin of pronotum from dark reddish brown to black; sculpture with dark reddish brown punctures on elytra and throughout anterior 1/6 of pronotum, in the latter the punctures are arranged in a triangular maculation. Teneral specimens can be completely or more homogeneously lightly pigmented, pale to light reddish brown. Rostrum ( Figs 2A View FIGURE 2 , 3 View FIGURE 3 A–B) very short, 0.4–0.5 mm, r/p = 0.5–0.6, dorsally arcuate; strongly angulate-tumescent near basal 2/ 5 (apparently positively allometric); tumescence distinctively polished dorsally; with one evident tubercle (apparently positively allometric) near inner margin of each eye; dorsolateral carina irregularly rugulose (apparently positively allometric); broad, apically slightly enlarged; dorsal impression (near base) or rugulosity indistinct; antennal insertion near apical 2/5. Eye ( Fig 3 View FIGURE 3 A–B) subcircular, interommatidial microsetae only on central region of eye; interocular distance slightly narrower than apex of rostrum. Head ( Figs 2A View FIGURE 2 , 3 View FIGURE 3 A–B) evenly slightly convex, without dorsal elevation or projections; setation sparse, short and recurvate; setae longer along dorsal margin of each eye. Pronotum ( Figs 2A View FIGURE 2 , 3C View FIGURE 3 ) expanded (apparently positively allometric), but not globular (see Franz 2008, Fig. 1 View FIGURE 1 ), anterolaterally not impressed. Prosternum ( Figs 2A View FIGURE 2 , 3D View FIGURE 3 ) with procoxal cavities separated by a narrow septum; with dark brown transversal stripe in front of each procoxa. Elytra ( Figs 2A View FIGURE 2 , 4D View FIGURE 4 ) posteriorly not attenuate; anterior elevation indistinct; striae narrower than intervals; only stria I evidently sulcate, remaining striae very shallow and similar to elytral intervals; striae and intervals with one row of dark reddish brown subequal circular punctures, evidently separated. Abdominal ventrites ( Figs 2A View FIGURE 2 , 4E View FIGURE 4 ): II shorter than III+IV; V with posterior margin sinuous, medially projected. Abdominal tergites ( Figs 4 View FIGURE 4 A–C) convex, with eight visible tergites; lateral sclerites absent; spiracular sclerites I–VI distinguished from the median one, very weakly sclerotized, almost membranous, covered by spiculate patch (posteriorly-directed); spiracular sclerite VII fused with the median one; median sclerites complete (no median fissure), weakly sclerotized (especially I–II) being VII–VIII more sclerotized; setation on median sclerites (dorsal surface): I–II covered by minute posteriorly-directed microtrichia (lacking spiculate patches), III with only lateral spiculate patches (laterally-directed), IV–VI with lateral and median spiculate patches (posteriorly-directed), IV–VII with a fringe of setae on posterior margin; tergite VII ( Fig. 4C View FIGURE 4 ) trapezoidal in dorsal view, concave on posterior margin, covered by trifid fan-like setae, no plectra; tergite VIII ( Fig. 4C View FIGURE 4 ) subtrapezoidal, posterior margin rounded, covered by trifid fan-like setae become longer unifid setae on apical half. Sternum VIII ( Fig. 5A View FIGURE 5 ) (each sclerite) triangular, deflexed downwards on outer 1/3, with eight large unifid setae on inner margin. Sternum IX (spiculum gastrale) ( Fig. 5B View FIGURE 5 ) distinctly shorter than aedeagus (0.6–0.7); basal plate V-shaped; apodeme similar throughout, apically light sclerotized. Tegmen ( Figs 5 View FIGURE 5 C–D) 0.3–0.4 as long as aedeagus; basal piece Y-shaped (basal plate reduced); tegminal apodeme deflexed apically. Aedeagus ( Figs 5 View FIGURE 5 D–H) l/w = 4.2–5.1, widest near middle; each side with two (instead of only one, see Franz 2008: 6) inner separately arched rami projecting beyond dorsal margin along subapical region; lateral margins evidently sinuous; apex acutely projected and deflexed; basidorsal margin evidently triangular; basiventral margin rounded; endophallus with one basal plicate barrel-shaped sclerite; endophallus ventrally covered by large setae on apical ½ and dorsally (in the region of the ostium) with a pair of subapical plicate sclerotizations, each sclerotization large, triangular, deflexed apically and undulate on inner margin; ostium large. Aedeagal apodemes 0.8 as long as aedeagus.
Female ( Figs 2B View FIGURE 2 , 6–7 View FIGURE 6 View FIGURE 7 ). Length 1.6–2.1 mm, width 0.7–0.9 mm, l/w = 2.2–2.6. Integument ( Fig. 2B View FIGURE 2 ) homogeneously light brown, slightly darker on head, rostrum and anterior margin of pronotum; darker punctures on elytra and on triangular maculation of pronotum lighter than male; teneral specimens can be completely or more homogeneously lightly pigmented. Rostrum ( Figs 2B View FIGURE 2 , 6A View FIGURE 6 ) 0.4–0.5 mm, shorter than pronotum r/p = 0.7–0.8, slightly arcuate, evenly convex, apically slightly expanded; lacking carina, tumescence, expansion or tubercles; outline contiguous with head; antennal insertion near middle. Pronotum ( Fig. 2B View FIGURE 2 ) slightly expanded, without apparent allometry (see also Franz 2008: 4; 6). Prosternum ( Figs 2B View FIGURE 2 , 6B View FIGURE 6 ): procoxal cavities as male; transversal impression in front of each procoxa much lighter than in male. Abdominal ventrites ( Figs 2B View FIGURE 2 , 6D View FIGURE 6 ): II longer than III+IV; V with posterior margin truncate. Abdominal tergites convex, with seven visible tergites; tergite VII ( Fig. 6C View FIGURE 6 ) triangular in dorsal view, without fringe of longer microtrichia on posterior margin. Tergum VIII ( Fig. 7A View FIGURE 7 ) concealed in the abdomen (not visible without dissection); 0.3x as long as sternum VIII, covered only by simple (unifid) setae, lateral margins acutely projected antero-ventrally; posterior margin rounded. Sternum VIII ( Fig. 7C View FIGURE 7 ) lamina pentagonal-shaped, furcal arms weakly sclerotized, connected only by membranous, not closed, covered by 19–23 subequal short setae on distal 1/4, apices no longer setae; apodeme arcuate ventrally. Spermatheca ( Fig. 7D View FIGURE 7 ) C-shaped, deflexed; corpus swollen; apex of cornu acuminate and deflected, without projection on outer margin at point of deflection; collum and ramus narrowly separated and not protruded; gland large, covered by spinules. Coxite ( Fig. 7B View FIGURE 7 ) shorter than sternum VIII, bearing small simple setae, slightly narrowed and curved posteriorly, inner margin sinuous; styli ( Fig. 7B View FIGURE 7 ) apically with three long simple setae.
Type material. Holotype male deposited in MPEG: “Brasil-PA-Belém\ Parque Estadual do Utinga \ 12-VIII-2013 \ R.M. Valente Col. [label 1], Em inflorescência de\ Evodianthus funifer \7:40 horas\Amostra 03”[label 2]. Allotype female deposited in MPEG: same as holotype . Paratypes: same as holotype (22♂ (5 dissected), 22♀ (6 dissected), UFPA) ; “ 12-VIII-2013, 7:00 horas, Amostra 01” (1♂, 1♀, UFPA) ; “ 12-VIII-2013, 7:30 horas, Amostra 02” (2♂, 2♀, MZUSP) ; “ 14-VIII-2013, 6:30 horas, Amostra 06” (2♂, 8♀, MPEG) ; “ 14-VIII-2 0 13, 6:40 horas, Amostra 07” (2♂, 2♀ (1 dissected), UFPA) ; “ 14-VIII-2013, 7:40 horas, Amostra 09” (2♂, 2♀, AMNH) ; “ 16-VIII-2013, 6:20 horas, Amostra 10” (2♂, 2♀, CMNC) ; “ 16-VIII-2013, 6:45 horas, Amostra 11” (3♂, 7♀ (1 dissected), UFPA) ; “ 20-VIII-2013, 6:55 horas, Amostra 14” (2♂, 4♀, MPEG) ; “ 20-VIII-2013, 9:12 horas, Amostra 18” (2♂, 2♀, MCZ) ; “ 20-VIII-2 0 13, 9:20 horas, Amostra 19” (3♂ (1 dissected), 2♀, MPEG) ; “ 22-VIII-2013, 6:30 horas, Amostra 21” (1♂, 1♀, MPEG) ; “ 22-VIII-2013, 6:50 horas, Amostra 22” (8♂, 9♀, UFPA) ; “ 22-VIII-2013, 7:06 horas, Amostra 23” (2♂ (1 dissected), 2♀, UFPA) ; “ 22-VIII-2013, 7:36 horas, Amostra 24” (4♂, 2♀, MPEG) ; “ 22- VIII-2013, 7:56 horas, Amostra 25” (2♂, 1♀ (1 dissected), UFPA) ; “ 22-VIII-2 0 13, 8:50 horas, Amostra 26” (2♂, 2♀); “ 23-VIII-2013, 6:30 horas, Amostra 27” (3♂, 4♀, UFPA) ; “ 23-VIII-2013, 6:49 horas, Amostra 28” (2♂ (1 dissected), UFPA) ; “ 23-VIII-2013, 7:05 horas, Amostra 29” (2♂, 2♀); “ 27-VIII-2013, 7:00 horas, Amostra 30” (2♂, 2♀ (1 dissected), UFPA) ; “ 27-VIII-2 0 13, 7:15 horas, Amostra 31” (2♂ (1 dissected), 2♀); “ 27-VIII-2013, 7:45 horas, Amostra 33” (1♂, 2♀, MPEG) ; “ 27-VIII-2013, 7:50 horas, Amostra 34” (1♂, UFPA) ; “ 27-VIII-2013, 7:55 horas, Amostra 35” (1♂, UFPA) ; “ 28-VIII-2013, 7:29 horas, Amostra 42” (4♂, 5♀, UFPA) ; “ 28-VIII-2013, 7:54 horas, Amostra 44” (3♂, 5♀, UFPA) ; “ 28-VIII-2013, 8:18 horas, Amostra 46” (2♂, 2♀, MPEG) ; “ 28-VIII-2013, 8:50 horas, Amostra 49” (2♂, 2♀, UFPA) ; “Brasil-PA-Benevides \ 10-VIII-2013 \ R. M. Valente Col. [label 1], Em inflorescência de\ Evodianthus funifer \6:00–9:00 horas\”[label 2] (5♂, 11♀, UFPA) .
Etymology. This species is named after Dr. Inocêncio de Sousa Gorayeb, who mentored the senior author, and for his friendship and dedication to the study of Neotropical insects and to training students in entomology in the Amazon. The name is a noun in genitive case.
Natural History. Cotithene gorayebi has been collected on inflorescences of Evodianthus funifer ( Cyclanthaceae, Carludovicoideae ) at two disjunct localities in the state of Pará, Brazil; i.e., Benevides (1°21’53.28”S 48°14’28.17”W) and Belém (Parque Estadual do Utinga, 1°25’S 48°25’W), both in Amazon lowland rainforest. All specimens were collected in August 2013, from 6:00 AM to 10:00 AM, at elevations from 23 to 37 m above sea level. In both localities, the annual precipitation average is about 3,000 mm also the annual mean relative humidity (87%, monthly mean from 85% to 95%) and annual mean air temperature (26°C, minimum temperatures above 18 °C) vary little throughout the year ( INMET 2015). The climate is tropical rainforest (Af type, Köppen-Geiger’s Climate classification) which is characterized by average precipitation of at least 60 mm in every month so without a true dry season but with a wetter season from December to May.
Evodianthus funifer View in CoL is a root-climbing liana widely distributed in the Neotropical region, and it is the only known species of Evodianthus View in CoL ( Harling 1958, Leal & Forzza 2012). Among species of Cotithene, only Cotithene gorayebi and C. melanoptera (the only previously known South American species of Cotithene) are known in association with inflorescences of E. funifer View in CoL , but collected from distant and ecologically different regions. Cotithene gorayebi has been recorded from Amazon lowland rainforest whereas C. melanoptera has been recorded in Merida and Aragua, both in a region of Venezuela far from Amazonian forest. Merida and Aragua are located at elevations from 1200 to 1600 m above sea level ( Franz 2008) and covered by a mosaic of savannas, deciduous forests and cloud forests (see Huber & Oliveira-Miranda 2010). The climate is humid subtropical to equatorial arid (Köppen-Geiger’s Climate classification). Moreover, the localities belong to different biogeographic domains: Pará ( Brazil) belong to the Boreal Brazilian Domain while Merida and Aragua ( Venezuela) belong to the Pacific Domain, both comprised in the Brazilian sub-region, Neotropical Region (see Morrone 2014). Cotithene gorayebi is the first Amazonian representative of the genus.
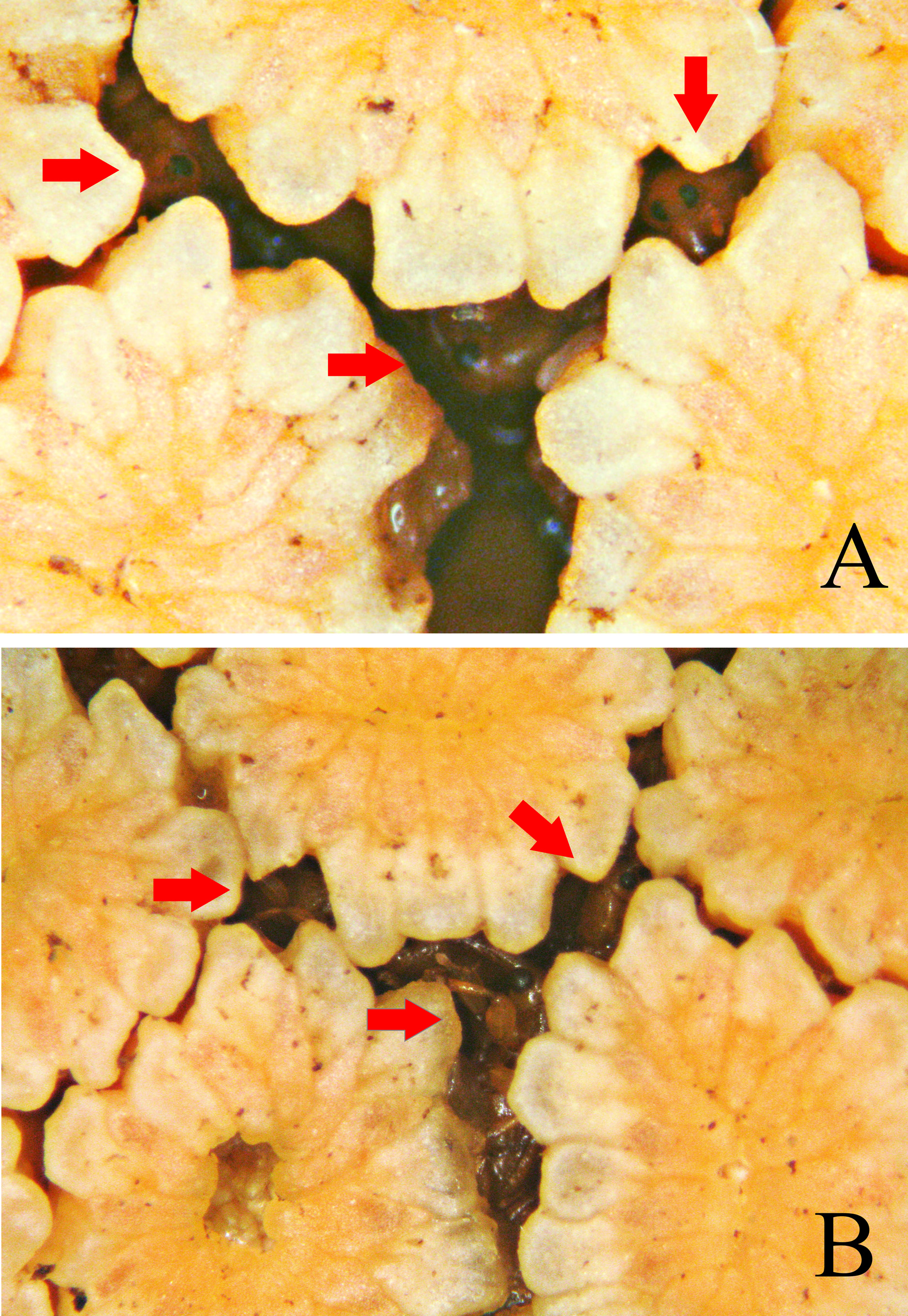
In the observations conducted at Parque Estadual do Utinga (Belém), E. funifer View in CoL behaved as a monoecious species with protogynous inflorescences (pistillate phase precedes staminate phase, see also Harling 1958, Gottsberger 1991). The pistillate phase ( Figs 8 View FIGURE 8 B–C) was short (about one day) and was recognized by droplets of exudate on pistillate flowers, stretched staminodes emitting a strong scent, and by staminate flower buds much more prominent and covering the pistillate flowers leaving just a small space above the stigmas. The staminate phase ( Fig. 8D View FIGURE 8 ) was very short (few hours) and recognized by dried stigmas, deteriorated staminodes and the opening of the perianth and anthers. One day before the pistillate phase, inflorescences began the phase prior to anthesis with open spathes but staminodes folded and odorless ( Fig. 8A View FIGURE 8 ).
Derelomini pollinators of cyclanths are attracted to the inflorescences by the scent emitted by the staminodes during the anthesis, especially during the pistillate phase ( Harling 1958; Gottsberger 1990, 1991; Eriksson 1994a; Franz 2004, 2007; Franz & O’Brien 2001a, 2001b). Here, the vast majority (92%) of adults of the new species were collected during the pistillate phase (more attractive to pollinators), a mean of 37 adults (range 16–126) per inflorescence (N=25). In the staminate phase, the sampling effort was conducted at 6:00 AM so after to the departure of pollinators (see Gottsberger 1991, Franz 2007). In this phase we recorded only 8% of all adults collected with mean of 9 adults (range 1–36) per inflorescence (N=15). However, inflorescences with at least scarce staminodes/pollen recorded 20– 36 adults of Cotithene gorayebi per inflorescence while those with fallen staminodes/released pollen (less attractive to pollinators) recorded only one to three adults.
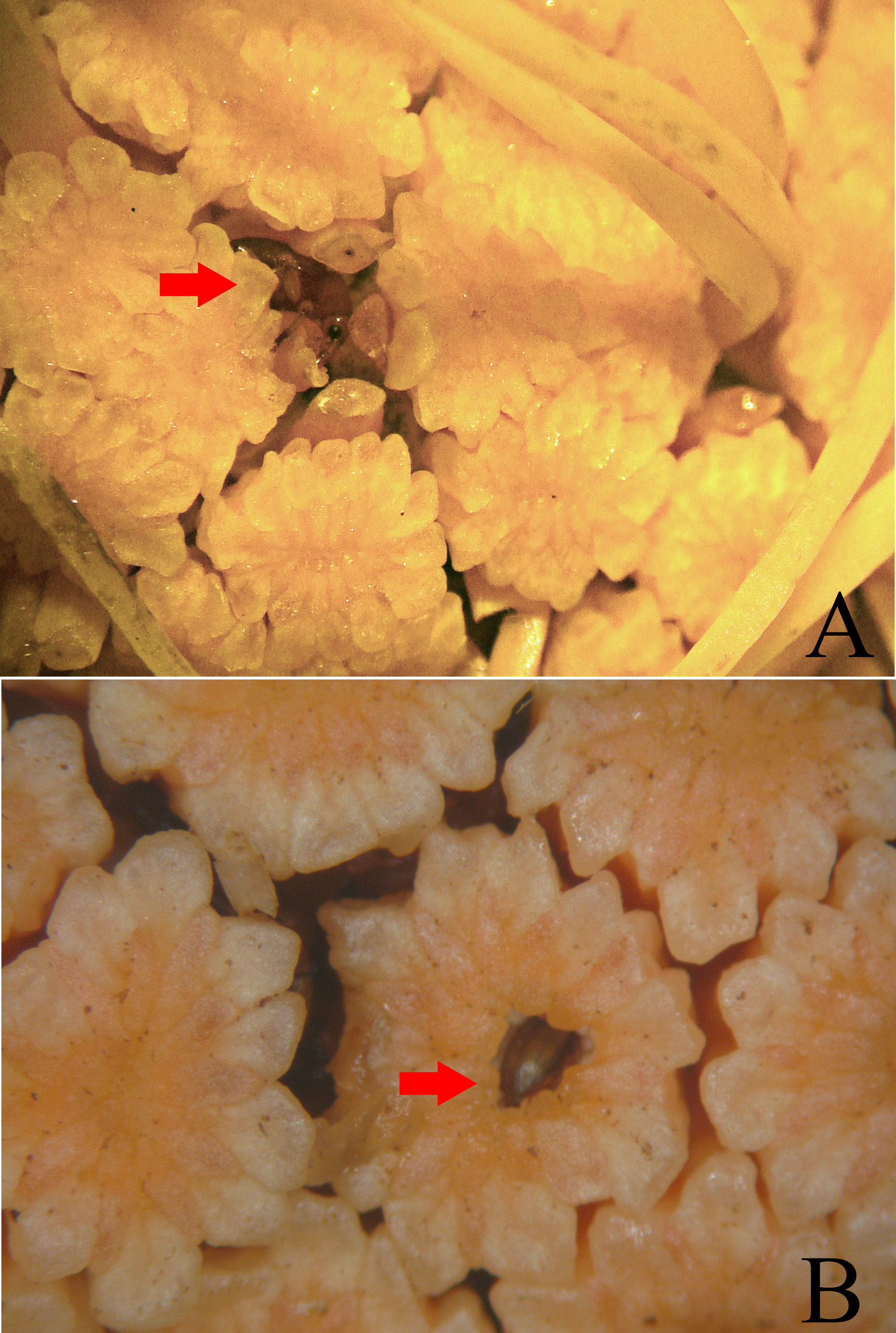
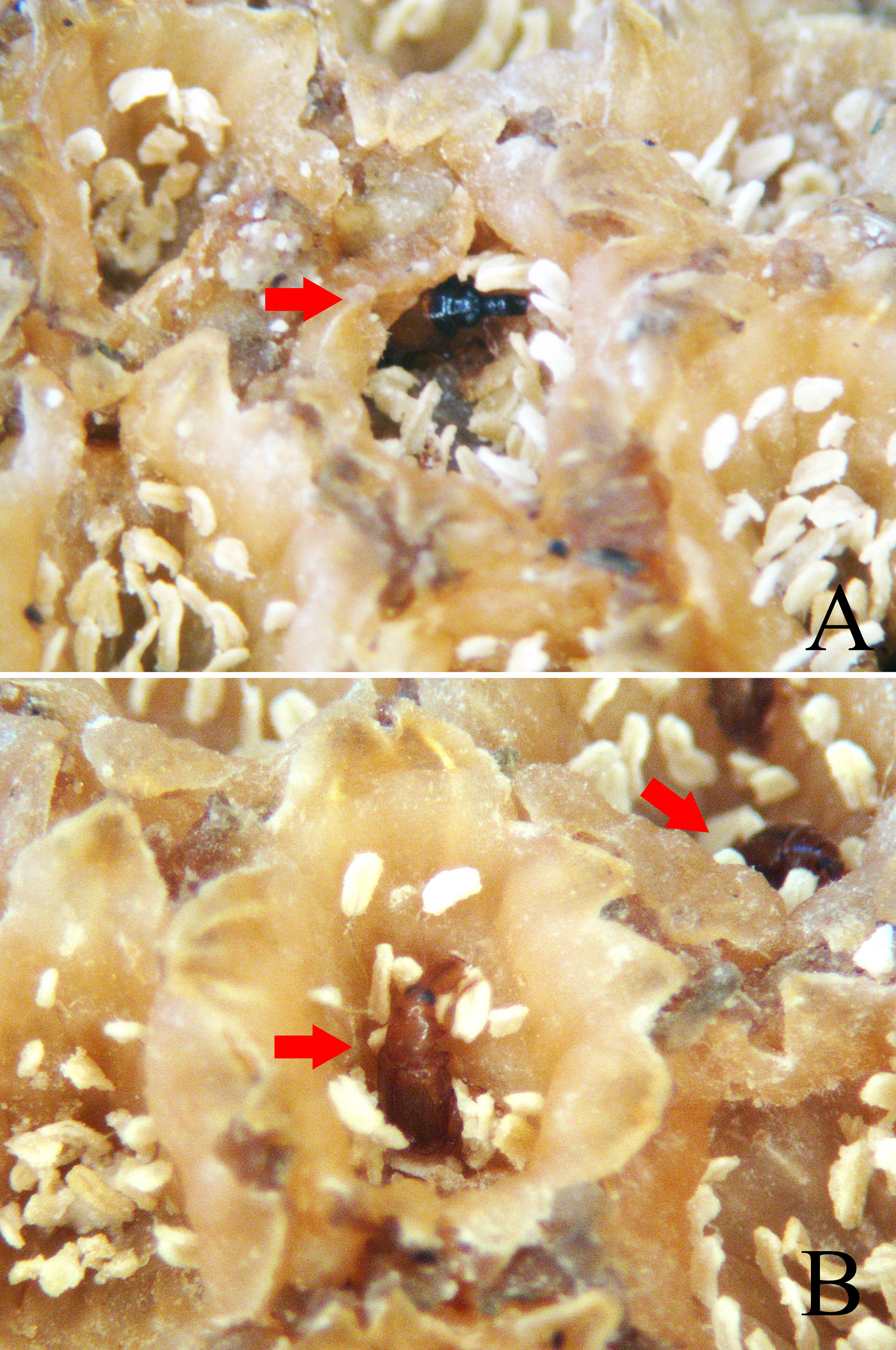
In the pistillate phase of E. funifer View in CoL , adults of Cotithene gorayebi were observed arriving in flight at the inflorescence with the first day-light (about 6:00 AM in August 2013). They immediately began to move towards the inner spaces of the inflorescences where the pistillate flowers are located and between the closed staminate flowers ( Fig. 9 View FIGURE 9 ). At these spaces, they fed on the base of staminodes ( Fig. 10A View FIGURE 10 ) and on the exudate present on pistillate flowers. Adults are small enough to reach the space where the stigma of female flowers are located and frequently touch the stigmas during the pistillate phase. Some adults chewed the perianth of closed staminate flowers to access the anthers ( Fig. 10B View FIGURE 10 ), and males were observed using the head and rostrum in male-male conflicts in the vicinity of the females. In the staminate phase, adults were already occupying spaces between staminate flowers ( Fig. 11 View FIGURE 11 ) before the observation effort (6:00–10:00 AM). These behaviors, together with their abundance (see above), suggests that adults of Cotithene gorayebi probably remain in the spaces between the closed staminate flowers until the staminate phase. They leave the inflorescences shortly after pollen has been released from anthers, often before the first day-light, as observed in other Derelomini pollinators of cyclanths ( Gottsberger 1990, 1991, Ericksson 1994a, Franz 2004, 2007). Finally, we observed pollen grains on the body of adults of Cotithene gorayebi , especially on the prosternum.
Previously known species of Cotithene are considered as non-pollinators of Cyclanthaceae because adults are scarcely abundant on inflorescences (typically 1–2 individuals per inflorescence) and present behavior not well synchronized with anthesis phases. Moreover, larvae develop into the rachis of inflorescence and can lead infructescence abortion ( Franz 2007, 2008). Here, we did not observe larvae of Cotithene gorayebi but adults were abundant on inflorescences of E. funifer (mean of 24 adults, N=40) and behaved synchronously with the phases of anthesis, as do other Derelomini pollinators of cyclanths. Behavioral patterns, in addition to the small size and high abundance of adults, strongly suggests that Cotithene gorayebi may be the first species of Cotithene described as playing a role as a pollinator of Cyclanthaceae . If larvae in this species also cause the abortion of a fraction of intructescences, Cotithene gorayebi is an additional example of a specialized brood pollinator that destroys seeds, a relatively rare kind of interaction between insects and plants ( Hembry and Althoff 2016), which only recently has been recognized in weevils ( De Medeiros et al. 2014, Nunes et al. 2018).
Remarks: Franz (2008: 5) states that species of Cotithene have a subapical foveola on the inner margin of procoxa. In stereomicroscopic view we also observed this feature in Cotithene gorayebi . Under scanning electron microscopy, we found that this foveola includes an internal circular projection ( Figs 12 View FIGURE 12 A–B).
The mouthparts ( Figs 12 View FIGURE 12 C–G) of Cotithene gorayebi are in accordance with Franz (2008: 4, Fig. 4 View FIGURE 4 ) but scanning electron microscopy also reveals that the new species bears a subapical obtuse projection on the inner tooth of the mandible ( Fig. 12D View FIGURE 12 ). Moreover, the maxilla ( Figs 12C View FIGURE 12 , E–F) bears six lacinial teeth [vs. 4–6 in the genus, see Franz 2008: 4, Fig. 4B View FIGURE 4 ] and the maxillary palps are 2-segmented (as in remaining Cotithene species) but maxillary palpomere II bears three elongate sensillae along the outer margin and two very small setae on the inner margin [both characters described for the first time to Cotithene]. Finally, the prementum of the labium ( Figs 12C, G View FIGURE 12 ) is rectangular [vs. subquadrate, see Franz 2008: 4, Fig. 4C View FIGURE 4 ] and its large setae along lateral margin is inserted subapically [vs. medially, see Franz 2008: 4, Fig. 4C View FIGURE 4 ]. Also note the evident apical triangular acute projection on the outer margin of labial palpomere II ( Fig. 12G View FIGURE 12 ) which is synapomorphic for the Cyclanthura–Cotithene clade ( Franz 2006, 2008).







| MPEG |
Brazil, Para, Belem, Museu Paraense Emilio Goeldi |
| MZUSP |
MZUSP |
| AMNH |
USA, New York, New York, American Museum of Natural History |
| CMNC |
Canada, Ottawa, Canadian Museum of Nature |
| MCZ |
USA, Massachusetts, Cambridge, Harvard University, Museum of Comparative Zoology |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
| AMNH |
American Museum of Natural History |
| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |