
Comanthus
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4268.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:0742D287-B82C-4014-A6AC-C357F259D5D7 |
|
DOI |
https://doi.org/10.5281/zenodo.6009172 |
|
persistent identifier |
https://treatment.plazi.org/id/039EDF70-FFA0-A141-FF66-DF0512BDFE55 |
|
treatment provided by |
Plazi |
|
scientific name |
Comanthus |
| status |
|
Comanthus AH Clark, 1908d
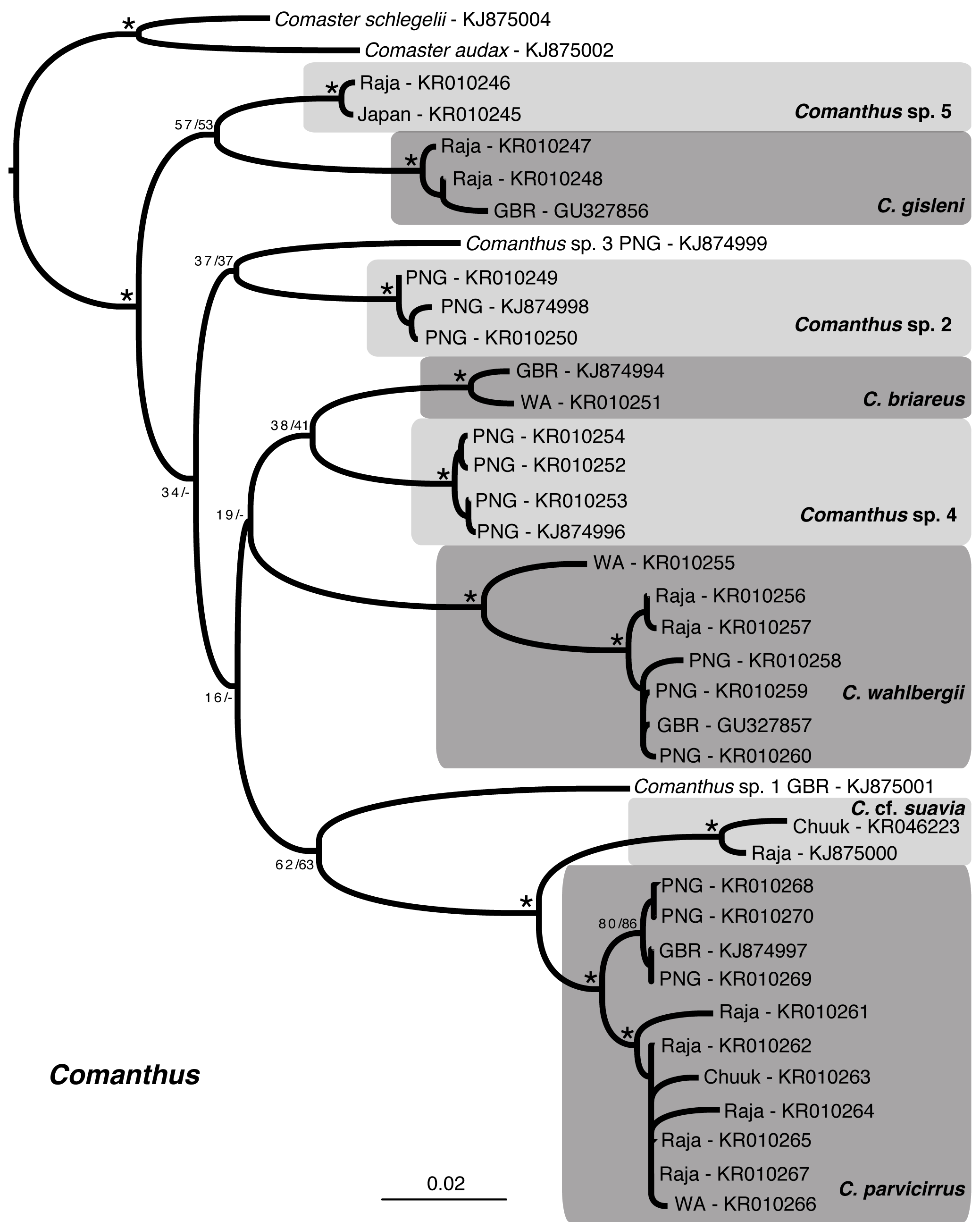
Table 1; Figs. 15–16 View FIGURE 15 View FIGURE 16
Type species. Alecto parvicirrus Müller, 1841 .
Other included taxa (9). Alecto wahlbergii Müller, 1843 ; Antedon briareus Bell, 1882 ; Comantheria weberi AH Clark, 1912a; Comanthus gisleni Rowe, Hoggett, Birtles & Vail, 1986 ; Comanthus kumi Fujita & Obuchi, 2012 ; Comanthus suavia Rowe, Hoggett, Birtles & Vail, 1986 ; Comaster imbricatus AH Clark, 1908e; Comaster taviana AH Clark, 1911e; Phanogenia delicata AH Clark, 1909f.
Diagnosis. Mouth excentric in fully developed individuals; up to 75 arms; centrodorsal circular to pentagonal; cirri present in one row or absent ( Fig. 15 View FIGURE 15 B, D–F, H–I, K–L); IBr2 united by synarthry; IIBr and subsequent brachitaxes 2 or 4(3+4); first syzygy at 3+4 on all undivided arms; distal intersyzygial interval 4; distalmost pinnule comb at intervals sometimes reaching near arm tip; comb terminating in large discrete tooth; proximal tooth transverse, saucer-like ( Fig. 15 View FIGURE 15 C); primary comb teeth confluent with exterior edge of pinnulars ( Fig. 15 View FIGURE 15 C); smaller secondary tooth present or not.
Distribution. Indo-western Pacific from northern Australia (Perth, WA, to Julian Rocks, NSW) west to the Red Sea and eastern Africa as far south as Cape of Good Hope, east to Kwajalein Atoll and Vanuatu, and north to Chiba Peninsula, Japan, and possibly the Korean Straits (AH Clark 1931; Kogo 1998; Rowe & Gates 1995; Zmarzly 1984). Depth range: 0–120 m (chiefly <50 m).
Molecular results. The sequenced specimens identified as Comanthus were recovered in multiple clades, with varying support ( Fig. 16 View FIGURE 16 ). All clades named in Fig. 16 View FIGURE 16 were assignable based on morphology and were recovered with interspecific uncorrected COI distances of at least 5%, with the following exceptions: 1) Comanthus sp. 5 and Comanthus gisleni (3.4–4.3%); 2) Comanthus cf. suavia and Comanthus parvicirrus (3.8–5.3%); 3) Comanthus sp. 2 and Comanthus sp. 3 (4.7%); 4) Comanthus sp. 5 and Comanthus sp. 2 (~4%); and 5) Comanthus sp. 4 and Comanthus briareus (4.1%). Intra specific distances (where available) were less than 1.3%, with the exception of Comanthus parvicirrus (<2.7%) and Comanthus wahlbergii (<3.4%). Five clades interspersed among those that could be named ( Fig. 16 View FIGURE 16 ) are arguably species-level taxa but were left unnamed owing to the dozens of available names that are currently treated as synonyms for C. wahlbergii and C. parvicirrus (e.g., AH Clark 1931; Rowe et al. 1986).
Remarks. It is not surprising that additional taxa likely exist within Comanthus , as the majority of current nominal species are widely distributed, include numerous synonyms, and some include different, probably habitatrelated, forms (e.g., Comanthus parvicirrus , Comanthus wahlbergii ). Distinguishing species of Comanthus remains difficult, as clear molecular boundaries for ‘species’ assignments are unclear.
SUBFAMILIAL ASSIGNMENT UNCERTAIN

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.