
Poecilimon nonveilleri, Ingrisch, Sigfrid & Pavićević, Dragan, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.197230 |
|
DOI |
https://doi.org/10.5281/zenodo.6204488 |
|
persistent identifier |
https://treatment.plazi.org/id/039D4825-F654-FFFA-FF12-FE25FD62FEC3 |
|
treatment provided by |
Plazi |
|
scientific name |
Poecilimon nonveilleri |
| status |
sp. nov. |
Poecilimon nonveilleri sp. n.
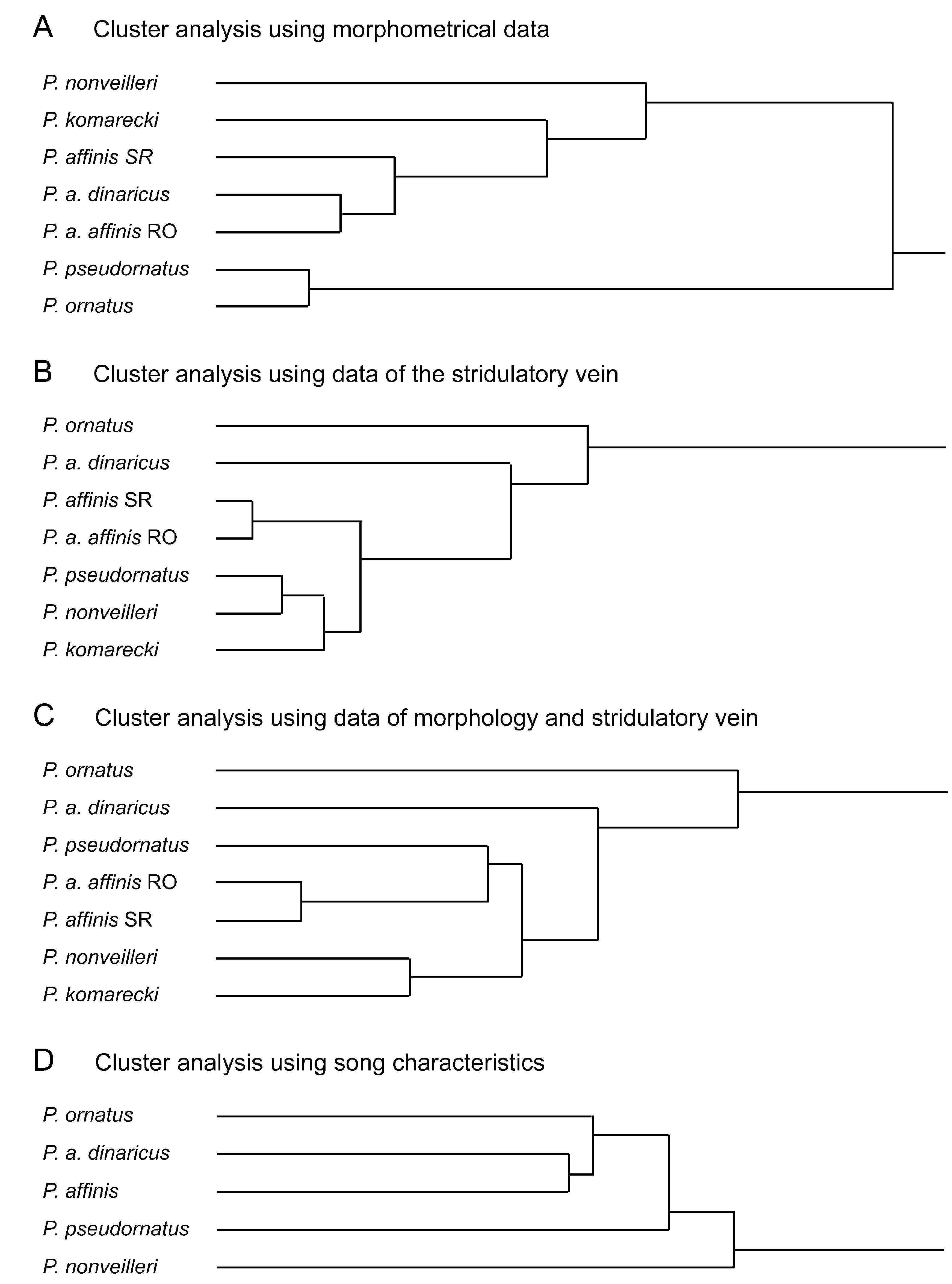
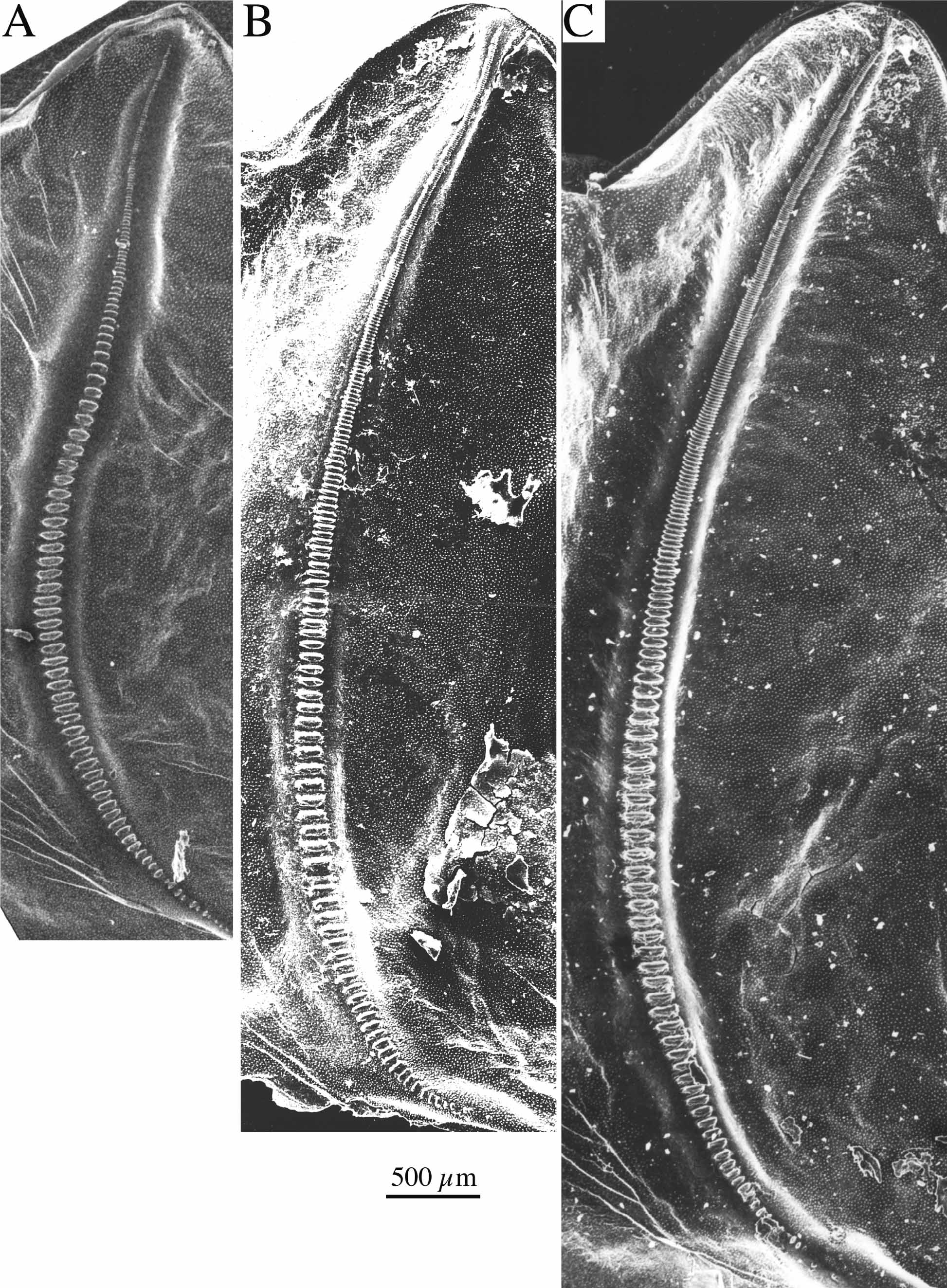
Figs. 6 View FIGURE 6 , 7 View FIGURE 7. A – H I –Q, 10B, 12; Tabs. 5–6 View TABLE 5 View TABLE 6 .
Holotype (male): Montenegro: Durmitor, Canyon of river Tara, Đurđevića Tara, 600m, 21.VII.1987, leg. S. Ingrisch, deposited in Muséum d'Histoire naturelle de Genève ( MHNG).
Paratypes: Montenegro, Durmitor: 7 3, 1 Ƥ, Canyon of river Tara, Đurđevića Tara, 600m, 22.VI.– 3.VIII.1985, leg. D. Pavićević ( CDPV); 4 3, 1Ƥ, Đurđevića Tara, 600m, 21.VII.1987, leg. D. Pavićević ( CDPV); 1 3, same locality, 28.VI.1988, leg. S. Ingrisch (CI); 1 3, do., 3.VII.1988; 2 3, do., 9.VIII.1989; 14 3, 2 Ƥ, same locality, 24.VI–26.VI.1990, leg. D. Pavićević ( CDPV); 1 Ƥ (allotype), do., 24.VI.1991 ( MHNG); 1 3, Bistrica-Tara, 25.VII.1987, leg. D. Pavićević ( CDPV); 2 3, same locality, 12.VIII.1989, leg. S. Ingrisch (CI); Škrke, 1730m, 9.VIII.1985, leg.A.Vujić ( CDPV); 1 3, Krecmani, 2000m, 4.VIII.1988, leg. D. Pavićević ( CDPV); 2 3, 3 Ƥ, Pirlitor, Pleća, 1000m, 2.VII.1990, leg. D. Pavićević ( CDPV); 2 3, Pirlitor, Selina, 1000–1100m, 5.VIII.1990, leg. Ingrisch & Pavićević (CI + CDPV); 2 Ƥ, Sušica Canyon, 1200m, 9.VIII.1985, leg. D. Pavićević ( CDPV); 7 3, 14 Ƥ, same locality, 28.VII.1988, leg. Ingrisch & Pavićević (CI + CDPV); 7 3, 4 Ƥ, Piva between Mratinje – Šćepan Polje, 500m, 29.7.1988, leg. Ingrisch & Pavićević (CI + CDPV); 2 3, Piva, SE Mratinje, 6.VIII.1989, leg. S. Ingrisch (CI); 2 3, Piva, SE Kruševo, 6.VIII.1989, leg. Ingrisch & Pavićević (CI + CDPV); 2 3, 1 Ƥ, Crkvičko Polje, 1100m, 13.VIII.1989, leg. Ingrisch & Pavićević (CI + CDPV); 2 3, 2 Ƥ, same locality ex ovo, leg. S. Ingrisch (CI);; 3 3, Gornja Grabovica-Gabelja, 1450m, 29.VII.1993, leg. D. Pavićević ( CDPV); 1 3, Grabovički do, 1700m, 29.VII.1993, leg. D. Pavićević ( CDPV); 1 3, 1 Ƥ, Bjelasica Mt., Biogradska Gora, 900m, 4.VII.1990, leg. D. Pavićević ( CDPV); 1 3, 2 Ƥ, do, 7.VII.1991.
Type locality. Tara Canyon (Durmitor, Montenegro), forest glades in mixed deciduous forest along the road between Tara bridge and Bistrica.
Measurements. see Tabs. 5–6 View TABLE 5 View TABLE 6 .
Diagnosis. The new species is similar to P. rumijae and P. affinis . From P. a f f i n i s it differs by the very narrow fastigium verticis, by the male stridulatory file and by stridulation. From P. rumijae it differs by the male pronotum being more strongly widening behind and leaving a greater part of the tegmen free. The stridulatory files of both species carry about the same number of teeth, but the file is shorter and the thus the teeth density higher in P. nonveilleri than in P. r u m i j a e. In the female the ovipositor is distinctly shorter in P. nonveilleri than in P. r u m i j a e. The differences to other species are outlined in the key and discussed above.
Description. A medium sized, rather stout species with very narrow fastigium verticis. Fastigium verticis distinctly narrower than scapus ( Tab. 5 View TABLE 5 , Figs. 7 View FIGURE 7. A – H M–N), furrowed above. Postfemora with 4–15 spinules on interior and exterior ventral margins.
Male. Pronotum ( Figs. 7 View FIGURE 7. A – H I–L) moderately to strongly raised in metazona; metazona with strongly diverging lateral margins; apex subtruncate to concave. Abbreviated tegmina covered by pronotum in circa basal third; stridulatory vein usually visible from above or above-behind. Stridulatory file ( Fig. 10 View FIGURE 10 B) with 200–250 teeth ( Tab. 6 View TABLE 6 ). Cerci ( Fig. 7 View FIGURE 7. A – H O–P) strongly curved before apex, more or less constricted before apical tooth. Subgenital plate narrowing behind middle with a faint median keel in apical half which is sometimes lacking; apex truncate or with a very shallow triangular excision; both apico-lateral corners often but not always pointed.
Female. Pronotum with dorsal margin substraight to distinctly raised in metazona; apex truncate to distinctly concave. Tegmina overlapping; hardly projecting behind pronotum; apex nearly truncate. Cerci very slightly curved; apex subacute to subobtuse. Subgenital plate triangular to almost semicircular, with a weak median keel; apex sometimes faintly protruding in middle. Ovipositor ( Fig. 7 View FIGURE 7. A – H Q) stout, apex dentate.
Coloration. Male. Light form: Green, indistinctly stippled with black. Antennae yellow. Pronotum with red or black or with red (externally) and black (internally) lateral bands of variable shape and length, present at least in metazona; discus in metazona partly yellow. Tegmina yellow, discus faintly infumate (brown). Abdominal tergites green with two yellowish white lateral bands and sometimes with up to four small black dots at the base of some tergites on both sides of the white bands.
Dark form: Yellowish brown to olivaceous, legs dark to reddish brown. Black dots more distinct than in the light form. Antennae infumate except at yellow base, but not distinctly annulated. Vertex black. Disc of pronotum with prozona almost completely black, metazona with a median and two lateral black bands which are fused along posterior margin. Tegmina yellow with disc distinctly infumate. Abdominal tergites with a median and two lateral black bands, the lateral bands interrupted before posterior margin of each tergite; last tergite black. In contrast to the dark form of P. pseudornatus , the epiproct is black, while the legs are dorsally not black.
Female. Light form: Uniformly green or less commonly yellowish brown when alive. Pronotum with red lateral stripes in metazona. Tegmina yellow.
Dark form: As in male.
The dark form appeared when specimens were bred from the egg in laboratory as with the other species mentioned above. It is thus not unlikely that it is induced by crowding during the nymphal development under breeding conditions. The brown colour variant was found in the Sušica canyon at a forest edge on litter.
Etymology. Named in honour of late Prof. Dr. Guido Nonveiller who initiated the faunistical investigations of the Durmitor mountain.
Distribution. With certainty only known from the canyons surrounding the Durmitor mountain, but extensive investigations will probably reveal a wider distribution in the Dinaric region. Upper altitudinal distribution is usually limited to about NN+ 1200m.
Stridulation ( Fig. 12 View FIGURE 12 ): As in P. pseudornatus , stridulation consists of monosyllables. However, the final, loud "zip" is distinctly longer ( Tab. 6 View TABLE 6 ). The intensity of the initial "sssss" is more gradually increased to the final pronounced part. Usually, males repeat a variable number of syllables in short intervals to form loose sequences. The main frequency is at 14–15 kHz.


| MHNG |
Museum d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.