
Sympagella multihexastera, Tabachnick & Menschenina, 2008
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1866.1.3 |
|
persistent identifier |
https://treatment.plazi.org/id/0398E958-487A-1576-FF41-09031033FD21 |
|
treatment provided by |
Felipe |
|
scientific name |
Sympagella multihexastera |
| status |
sp. nov. |
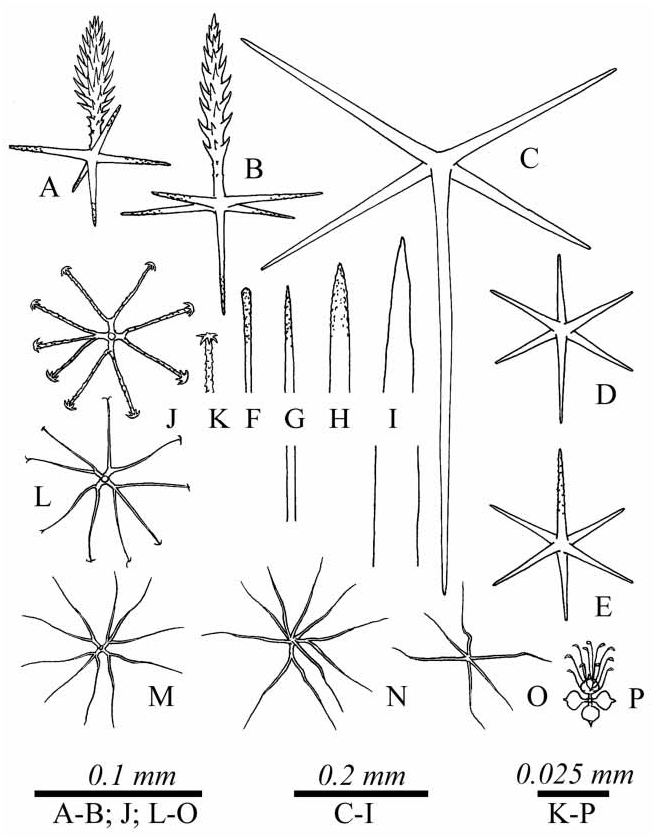
Sympagella multihexastera View in CoL sp. nov. ( Fig. 31 View FIGURE 31 , 32 View FIGURE 32 ; Tab 29)
Etymology. The name refers to the multitude of hexaster types characteristic of the spicule composition of this species.
Material. Holotype: NTM Z 0002592 View Materials —Sta. NWS0050, off Rowley Shoals, Northwest Shelf, Scampi trawl Grounds, 17 o 28’S 118 o 53’E, depth 405 m. GoogleMaps
Paratype: NTM Z 0002594 View Materials —Sta. NWS0052, off Rowley Shoals, Northwest Shelf, Scampi trawl Grounds, 17 o 28’S 118 o 53’E, depth 405 m. GoogleMaps
Description. Body: Both specimens are presented by lamellalike fragments, 5–8 mm in thickness.
Spicules: Choanosomal spicules are diactins and hexactins. The diactins have stout shafts, rarely with a widening in the middle and conically pointed or rounded outer ends, smooth or rough, they measure 1.5–2.9/ 0.01–0.13 mm. The hexactins have conically pointed outer ends, usually smooth, some hexactins have rough rays directed outside the body; the rays of these hexactins are 0.15–0.37/0.01–0.0.026 mm. Hypodermalia and probably hypoatrialia are pentactins with conically pointed, usually smooth outer ends, their tangential rays are 0.25–0.87 mm long, the ray directed inside the body is 0.3–1 mm, the diameter of these rays is 0.014 – 0.026 mm. Dermalia and atrialia are pinular hexactins with a spindlelike pinular ray, the other rays are conically pointed or rounded, usually rough, sometimes smooth. The pinular ray of dermal hexactins is 0.066– 0.143/ 0.009 mm, tangential rays are 0.041 –0.077 mm, the proximal ray is 0.041 –0.077 mm, the diameter of these rays is 0.002 –0.003 mm. The pinular ray of atrial hexactin is 0.092 –0.204 mm long, tangential rays are 0.041 –0.071 mm, the proximal ray is 0.051 –0.082 mm.
Microscleres: Microscleres are discohexasters, rarely onychohexasters, oxyhexasters, rarely oxyhemihexasters, oxyhexactins and strobiloplumicomes. The discohexasters, with 2, rarely 3 rays, are uncommon; they are 0.101 –0.105 mm in diameter with a primary rosette 0.008 –0.017 mm in diameter. The single onychohexaster found is 0.063 mm in diameter with a primary rosette 0.008 mm in diameter. The microscleres with oxyoidal secondary rays are 0.076 –0.113 mm in diameter with a primary rosette 0.008 –0.017 mm in diameter. The strobiloplumicomes are 0.029 –0.063 mm in diameter with a primary rosette 0.013 –0.025 mm in diameter.
Remarks. The new species ‘multihexastera’ is erected according to its specific combination of microscleres: discohexasters, onychohexasters, oxyhexasters, oxyhexactins and strobiloplumicomes (the latter are characteristic for all species of the genus). Probably the closest species with most similar microscleres is S. clavipinula Tabachnick and Levi, 2004 from off New Caledonia, but this species has no oxyhexactins, dermal pinular rays are clavate in shape and atrial pinular hexactins differ strongly from the dermal ones in shape and length of their pinular rays. Two other species which possess similar dermal and atrial spicules are S. johnsoni ( Schulze, 1887) (formerly Aulascus ) from off South Africa and S. gracile (Schulze, 1903) (formerly Calycosoma ) from the Indonesian Archipelago. The difference here is in the microsclere composition: S. johnsoni has (besides the characteristic strobiloplumicomes) discohexasters and onychohexasters, S. gracile has oxyhexasters and onychohexasters.
Discussion. Intraspecific variability and key to subspecies of Euplectella aspergillum . The result of this revision is the new definition of E. aspergillum as Euplectella having principalia of stauractins, notable fusion of the choanosomal skeleton, formation of ridges, microscleres as floricomes, oxyhexasters and their derivatives, including abnormal oxyoidal forms. Furthermore, usually also graphiocomes and sometimes sigmatocomes (which are probably young stages of floricomes) occur. The four subspecies are justified by their morphological differences as well as the fact that they inhabit distal geographical locations: E. aspergillum aspergillum Owen – off the Philippines; E. aspergillum regalis Schulze – Sea of Bengal; E. aspergillum indonesicum ssp. nov. – Indonesian Archipelago; E. aspergillum australicum ssp. nov. – off W Australia. Two subspecies are known from many specimens: ‘aspergillum’ and ‘australicum’, the other two only by single representatives. Their identification is based on the detailed investigation of oscularia (spicules located close to the lateral oscula) and of their microscleres: floricomes and oxyhexasters. No overlaps of important features for their identification was observed, therefore the identification key presented here seems justified. The other species of Euplectella , which show principalia of stauractins, require reexamination: It is very likely that E. oweni Herklots and Marshal, 1868 , E. marshalli Ijima, 1895 and E. curvistellata Ijima, 1901 should be synonymized; as well as E. imperialis Ijima, 1894 , E. timorensis, Ijima, 1927 and specimens of E. regalis from off S Australia ( Reiswig 1992).

| NTM |
Northern Territory Museum of Arts and Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |