
Hippasteria, NOZAWAI GOTO, 1914
|
publication ID |
https://doi.org/10.1111/zoj.12131 |
|
persistent identifier |
https://treatment.plazi.org/id/0395879A-1C1D-FFAD-FF51-FD6CB1186B2B |
|
treatment provided by |
Carolina (2021-06-01 14:20:17, last updated by Plazi 2023-11-02 19:05:34) |
|
scientific name |
Hippasteria |
| status |
|
HIPPASTERIA NOZAWAI GOTO, 1914 View in CoL
Goto, 1914: 344; Mah et al., 2010: 285, 286 (as H. nozawai ).
Type species
Hippasteria nozawai Goto, 1914 View in CoL , type specimen status unknown.
Comments
Based on the original description in Goto (1914), this species appears to share most similarity with H. leiopelta . Most of the characters, pedicellariae, marginal plate series, marginal plate surface granulation pattern, etc., are very similar with only the overall R: r body shape differing. The R: r ratio in H. nozawai is 2.0 vs. H. leiopelta , which has R: r = 1.5. It seems likely that the two species are closely related and/or possible synonyms. It is tentatively placed with H. leiopelta within the H. phrygiana cluster. No material of this species was available for examination.
Occurrence
Hokkaido, northern Japan (no depth information available).
Material examined
None.
HIPPASTERIA PHRYGIANA ( PARELIUS, 1768) View in CoL
( FIGS 7A–J View Figure 7 , 8A–J View Figure 8 )
Linck, 1733: 21 (as Pentaceros planus ).
Parelius, 1768: 425 [1770: 349] (as Asterias phrygiana ).
Lamarck, 1816: 555 [non A. equestris Retzius, 1805 ] (as Asterias equestris ).
Gray in Johnston, 1836: 146 (as Asterias johnstoni ).
Gray, 1840: 279, 1866: 9 (as Hippasteria europaea , Hippasteria johnstoni , Hippasteria cornuta ).
Gray, 1840: 279; 1866: 9; Perrier, 1875: 270 [1876: 65], Danielssen & Koren, 1882: 268; Sladen, 1883: 159; Perrier, 1888: 764; Sladen, 1889: 341; Koehler, 1909: 88, 1924: 179 (as Hippasteria plana ).
Forbes, 1841: 125 (as Goniaster equestris ).
Müller & Troschel, 1842: 52; Düben & Koren, 1846: 246 (as Astrogonium phrygianum ).
Forbes, 1843: 280 (as Goniaster abbensis ).
Barrett, 1857: 47 (as Astrogonium aculeatum ).
Norman, 1865: 128 (as Goniaster phrygiana ).
Dons, 1937: 17 (as Hippasteria [Euhippasteria] phrygiana and Hippasteria [Nehippasteria] insignis ).
Verrill, 1874: 413; 1885: 542, Ganong, 1893: 56; Grieg, 1895: 6; Verrill, 1895: 137, 1899: 148; Döderlein, 1900: 218; Hartlaub, 1900: 191; Ludwig, 1900: 457; Whiteaves, 1901: 50; Grieg, 1902: 21; Pearcey, 1902: 308; Simpson, 1903: 40; H.L. Clark, 1905: 1; Nordgaard, 1905: 160, 235; Grieg, 1907: 28, 32; Süssbach & Breckner, 1911: 215; Grieg, 1912: 6; 1913: 115; 1917: 8; 1921: 6; H.L. Clark, 1923: 270; 1926: 13; Mortensen, 1927: 88, 1932: 8; 1933: 245; Haubold, 1933: 200; Einarsson, 1948: 10; A.H. Clark, 1949: 373; Djakonov, 1950: 53; Blacker, 1957: 18, 45; Buchanan, 1966: 25; Wolff, 1968: 82, Walker, 1978: 361; Franz et al., 1981: 406, 415; O’Connor & Tyndall, 1986: 96; Clark & Downey, 1992: 247; Picton, 1993: 22, 23; Gulliksen et al., 1999: 73, 105; Anisimova & Cochrane, 2003: 117, 121; Moore et al., 2004: 246; Dilman, 2006: 181; Mah et al., 2010: 287; Mah, 2011: 27; Clark & Jewett, 2011: 48; Foltz et al., 2013: 1285 (as Hippasteria phrygiana ).
Bell, 1892: 76 (as Hippasterias phrygiana ).
Perrier, 1891: K128; A.M. Clark, 1962: 22 (key); Codoceo & Andrade, 1978: 156, 168; Branch et al., 1993: 44, 60; Larraín et al., 1999: 437; Stampanato & Jangoux, 2004: 7 (as Hippastera hyadesi ).
Verrill, 1909: 63; Fisher, 1911: 224; Goto, 1914: 349; Verrill, 1914: 301; Djakonov, 1950: 53 (1968: 45); Baranova, 1957: 162; Alton, 1966: 1702; Carey, 1972: 38, 39; Lambert, 1978a: 4, 14; Maluf, 1988: 34, 118; A.M. Clark, 1993: 259; Lambert, 2000: 64; Clark & Jewett, 2011: 53 (in key) (as Hippasteria spinosa ).
Fisher, 1911: 230 (as Hippasteria leiopelta armata ).
Fell, 1958: 11, pl. 1, figs A, G; 1959: 136, fig. 21; 1960: 61, pls. 2,3; 1962: 33; McKnight, 1967: 300; H.E.S. Clark, 1970: 3; A.M. Clark, 1993: 259; Rowe & Gates, 1995: 65; Koslow & Gowlett-Holmes, 1998: 44 (as Hippasteria trojana ).
Fisher, 1911: 226 (as Hippasteria spinosa kurilensis ).
Goto, 1914: 338; Hayashi, 1952: 147; 1973: 6, 15; Imaoka et al., 1990: 50; Mah, 1998a: 67; Fujikura et al., 2008: 272 (as Hippasteria imperialis ).
Clark, 1926: 13; A.M. Clark, 1952: 196; A.M. Clark & Courtman-Stock, 1976: 63 (as Hippasteria phrygiana strongylactis ).
Mortensen, 1933; A.M. Clark, 1952: 196; A.M. Clark & Courtman-Stock, 1976: 63 (as Hippasteria phrygiana capensis ).
Djakonov, 1950 (1968: 47), Gish, 2007; R.N. Clark & Jewett, 2011: 53 (in key) (as Hippasteria kurilensis ).
Djakonov, 1950: 22 (1968: 46) (as Hippasteria derjungini ).
Djakonov, 1950: 54 (1968: 45), 1952: 409 (as Hippasteria mammifera ).
Djakonov, 1950: 54 (1968: 45); 1952: 410 (as Hippasteria pedicellaris ).
Djakonov, 1950: 55 (1968: 47); 1952: 411; Baranova, 1957: 162 (as Hippasteria colossa ).
Bernasconi, 1961: 1 (as Hippasteria argentinensis ).
Djakonov, 1950: 22 (1968: 46) (as Hippasteria spinosa f. armata ).
Clark & Downey, 1992: 248 (as Hippasteria phrygiana argentinensis ).
R.N. Clark & Jewett, 2011:52 (as Hippasteria armata ).
R.N. Clark & Jewett, 2011:48 (as Hippasteria aleutica ).
Type specimens
Asterias phrygiana Parelius, 1768 . Clark & Downey (1992) speculated that the type may be in the MNHN collections, but not located. A search by one of the authors (C. M.) further concluded that the type could not be located in the MNHN collection. Type specimens for recently synonymized names are below. The status of Hippasteria species described by Goto (1914) remains unknown pending further search of Japanese type repositories. Although the British Columbia syntype of H. spinosa View in CoL is listed below, the Puget Sound syntype has not been located. Its status remains unknown. Djakonov’s Hippasteria View in CoL specimens are deposited at the Zoological Institute in St. Petersburg Russia (ZI − abbreviation by C.M.).
List of relevant type specimens from recent synonymy: H. aleutica : Holotype: LACM 2008-030.001; Paratypes USNM 1125124, E47392 View Materials ; CASIZ 180539; University of Alaska, Fairbanks INV 8140; NOAA Auke Bay Laboratory AB11-002; LACM 1997-221.001, 1997- 154.013, 1999-183.001, R.N. Clark personal collection (ten miscellaneous specimens). Hippasteria argentinensis : holotype, Argentine Museum of Natural Sciences 9069; H. colossa ZI holotype no. 1/27102; Hippasteria derjungi ZI holotype no. 1/27101; H. hyadesi holotype, MNHN EcAh 2840; H. imperialis type status unknown; H. mammifera ZI holotype no. 1/7103; H. pedicellaris ZI holotype no. 1/7104; H. phrygiana capensis : syntypes, Copenhagen Museum, H. phrygiana strongylactis , holotype, South African Museum A22531 View Materials , paratypes MCZ 2831, 2864; H. spinosa , syntype, National Museum of Natural Sciences, National Museum of Canada NMCICI900-2933; H. spinosa armata (= H. leiopelta armata ) USNM 27887; H. spinosa kurilensis (= H. kurilensis ), holotype USNM 27885, paratypes USNM 32103, 32106; H. trojana Holotype, Canterbury Museum, Christchurch.
Synonymy
Results from Foltz et al. (2013) listed several previously accepted species as synonyms of H. phrygiana ( Parelius, 1768) , the type name for species within Hippasteria ( Clark, 1993; Mah & Hansson, 2012). Sampled taxa that were shown to be synonyms of H. phrygiana by Foltz et al. (2013) include H. spinosa Verrill, 1909 , H. armata ( Fisher, 1911) , H. kurilensis Fisher, 1911 , H. armata Fisher, 1911 , and H. aleutica Clark & Jewett, 2011 . Foltz et al. (2013) further removed any doubt regarding synonymy of the Patagonian H. hyadesi Perrier, 1891 or the New Zealand H. trojana Fell, 1958 into H. phrygiana .
Several North Pacific Hippasteria species show few if any morphological differences from H. phrygiana and are almost certainly synonyms. In the North Pacific, this includes H. colossa Djakonov, 1950 , H. derjungini Djakonov, 1950 , Hippasteria mammilifera Djakonov, 1950 , and H. pedicellaris Djakonov, 1950 . R.N. Clark & Jewett (2011) raised H. armata to a species without any apparent justification. Inclusion of H. armata in Foltz et al. (2013) showed that it falls within the synonymy of H. phrygiana along with many other previously established North Pacific Hippasteria spp. Based on the size of specimens and comparisons of appropriately sized specimens of H. phrygiana with the H. armata morphotype, these were almost certainly small-sized individuals.
The results of Foltz et al. (2013) are consistent with historical accounts that have shown H. phrygiana to be either very similar to other species or to be nearly identical but separate because of the distance separating individuals. A.M. Clark (1962) indicated that the type locality outlined for H. hyadesi by Perrier (1891) and Koehler (1926) must be mistaken because the specimen was essentially indistinguishable from H. phrygiana (= H. plana ). Based on our identification of Hippasteria from this region there is no longer a reason to discount the type locality as mistaken. Before being synonymized by H.E.S. Clark & McKnight (2001), Fell (1958) described the South Pacific H. trojana and noted its similar appearance and close relationship to the North Atlantic H. phrygiana . The type description of the North Pacific H. spinosa by Verrill (1909: 63) describes it as ‘Very similar in form and size to H. phrygiana of the N. Atlantic...’ with further remarks on close affinity by Fisher (1911). Other synonyms in the Atlantic almost certainly include H. phrygiana argentinensis and H. phrygiana capensis although we did not include specimens from those regions ( Argentina and South Africa) in our analysis.
Accounts that have described subspecies in Hippasteria [e.g. H. phrygiana argentinensis and others outlined in Clark & Downey (1992)] could not be substantiated by consistent character differences (e.g. spine counts with continuous overlap, etc.). Geographical clusters amongst major H. phrygiana haplotypes were arranged for methodological necessity (see Foltz et al., 2013) and did not reflect any significant morphological differentiation amongst species. Sampling from some localities (e.g. Solomon Islands) was anecdotal and suggests more widespread occurrence may be present in poorly sampled areas, which may further elucidate the phylogeny in Figure 1 View Figure 1 .
Occurrence and bathymetry
North Pacific: Japan, Aleutian Islands, Alaska, west coast of North America, including Canada, Washington, Oregon, west to President Jackson Seamount, south to Southern California. South Pacific: New Zealand, Solomon Islands, Straits of Magellan-Chile. South Indian: Kerguelen Islands, Crozet, Marion and Bouvet Isles, South Africa. Atlantic: east coast of North America, south to Cape Cod. North-eastern Atlantic−North Sea: Kattegat, northern Scotland and northernmost Ireland, the UK. South Atlantic: off Argentina, west side of South Africa, Lambert’s Bay to Cape Point.
Collectively throughout its range, H. phrygiana occurs widely from subtidal to continental shelf/upper bathyal depths, 10–1405 m. However, note that regional bathymetric occurrence varies. H.E.S. Clark & McKnight (2001) noted that H. phrygiana in New Zealand waters occurs primarily at> 500 m depths, seldom occurring at shallower depths. By contrast, Lambert (2000) reported this species (as H. spinosa ) along the Pacific north-west coast of North America at depths ranging from 10– 512 m. Maluf (1988) listed the depth of H. spinosa as present between 49–1170 m. Clark & Downey (1992) recorded North Atlantic H. phrygiana as being present in as shallow as 20 m (to 457 m in the Baltimore Canyon), but occurring in the South Atlantic and South Indian Ocean at depths to 310 m. Dilman (2006) noted occurrence of H. phrygiana to the Reykjanes Ridge based on an earlier account by Einarsson (1948). Einarsson (1948) was checked but the reference to Hippasteria could not be verified.
Comments on variation
Variation in H. phrygiana is extensive and although we have attempted to synthesize the extent of this variation, the summary presented here may be imprecise where localized populations are concerned. It remains helpful to refer to regional monographs that offer comprehensive, if older accounts, of H. phrygiana . In the North Pacific, Fisher (1911) summarized H. phrygiana populations (as H. spinosa and others) along the Northwest Pacific coast. Clark & McKnight (2001: 54) pre- sented an overview of variation in H. phrygiana between the New Zealand and the Atlantic specimens. Clark & Downey (1992) provided a taxonomic summary of North Atlantic H. phrygiana . Accounts of high latitude members (e.g. as H. hyadesi ) are more scattered and not as comprehensive (e.g. Perrier, 1891; Bernasconi, 1963; Stampanato & Jangoux, 2004).
It was difficult to conclude if character variation observed within H. phrygiana could be definitively associated with any observed environmental parameter (i.e. depth, location). Variable characters across the species’ range, which have historically been used to distinguish species, included accessory granules, spines, and all aspects of pedicellariae, including size, shape, abundance, and distribution.
Some deeper-water individuals (e.g. USNM 1215324 About USNM from 1405 m depth) displayed finer and more acutely tipped spination with relatively small body size, whereas others from more distant regions (e.g. IE IE- 2007- 1311 in the Solomon Islands) from comparable depths (722 m) were large with robust spines. Fewer samples are available from these settings, and regional vs. bathymetric variation are likely to be key factors .
An unusual aspect of the observed variation of H. phrygiana across its geographical range was the differing R: r ratios (i.e. the degree to which the arms are more distinct relative to the arms) at comparable sizes for different populations. For example, Figure 7G, H View Figure 7 shows two individuals at approximately identical sizes ( R = ∼4.4/4.5). One from the North Atlantic ( USNM E46619 View Materials ) shows a much stronger stellate shape vs. one from the Crozet Islands (MNHN EcAs 10683), which has a larger disk and broader arms. Environmental factors affecting growth, such as the amount of nutrients and degree of available calcium carbonate, would presumably be significant factors in affecting ontogenetic variation amongst differing populations within this species.
Description
Body in adults, strongly stellate, disk broad, thick ( Fig. 7A, D View Figure 7 ). Arms triangular, distinct, short (at R = ∼ 0.5 cm) to long, triangular (in adults, R > ∼ 2 cm). Inter-radial arcs strongly curved to straight. R: r = 2.0– 1.75 (adults) to 1.0 (small individuals). Large individuals R = ∼15.0 cm.
Abactinal surface convex, surface often tumescent. Individual plates round to oval to irregular in outline. Smaller, more oval- to irregular-sized plates occurring distally adjacent to contact with superomarginal series. Larger, more circular plates occurring more proximally especially near disk. Primary circlet and carinal series variably distinct to indistinct. More evident in New Zealand, Chilean/sub-Antarctic individuals, less clearly evident in North Atlantic and North Pacific individuals. Abactinal plates all bordered by accessory granules, two to 30, which vary in size and shape across different populations. These range from small, beadlike granules (<0.5 mm) or sharp, granular spinelets to larger, quadrate, blocky granules (1–2 mm) sometimes with no apparent regional distinction. For example, some New Zealand specimens show both the blocky granules ( USNM E13768) to small, bead-like granules ( NIWA 38153). More blocky granules are more closely arranged vs. the smaller accessory granules, which are more spread apart. Abactinal plates weakly convex and plate surfaces are bare except for spine or pedicellariae. Exceptionally plates lack both spines and pedicellariae and are completely smooth and bare. Shallow but well-developed fasciolar grooves present around abactinal plates extending to marginal and actinal surfaces. Madreporite convex, polygonal in shape, prominent (5–7 mm in diameter) with distinct grooves.
A large (often about 1.0 mm in diameter and up to 5 mm in length), single spine, conical to cylindrical and more blunt-tipped in shape, is present on a majority of abactinal plates but some variable individuals do show more pedicellariae relative to spines. As with the plates, largest spines are present proximally on the disk, becoming shorter and more tubercle like distally, adjacent to the superomarginal contact. Some New Zealand and Chilean/Southern Hemisphere individuals seem to display longer, sharper abactinal spines compared to their more northern counterparts, which have shorter, blunter spines.
Abactinal pedicellariae bivalve (exceptionally trivalve) vary in size and shape but are abundant on abactinal surface, ranging in shape from large-sized with more elongate/bivalve valves (Atlantic) to more elongate with rounded valves bearing weakly expressed but jagged teeth. Atlantic individuals observed have large, wide (up to 3.0 mm length at R = 10.6 cm) valves ( Fig. 7B View Figure 7 ). These individuals with smooth, elongate bivalve pedicellariae with smooth valves present on those plates lacking spines, roughly occurring one to every five plates with a spine. Extensive variation is observed in North Pacific, especially Aleutian Island, individuals, with shorter, more rounded, curved more elongate valves (up to 2.0 mm length at R = 11.3 cm) with some individuals bearing pedicellariae that outnumber spines on abactinal disk plates ( Fig. 7J View Figure 7 ). The Solomon Islands specimen ( IE- 2007-1311) has pedicellariae significantly smaller than those observed on the other specimens. South Pacific ( New Zealand) individuals have smaller pedicellariae overall with several bearing more elongate (defined as length of valve> width of valve) valves each bearing two to four weakly jagged teeth. Larger bivalve pedicellariae with smooth valves have also been observed.
Marginal plates broadly quadrate to polygonal in shape (e.g. Fig. 7C, E, I View Figure 7 ) but variable in length vs. width, ranging from more elongate to more equidimensional (L = W). Superomarginals correspond 1:1 with inferomarginals most strongly inter-radially, becoming more offset distally. Number of marginals is variable but ranges (measured arm-tip to arm-tip) from 32 to 52. Marginal plate surface is bare in most save for large spines and pedicellariae with the exception of the individual specimen from the Solomon Islands ( IE- 2007-1311), which has several concentric rows of granules covering the marginal plate surface up to the spine base on each plate. However, even in that specimen, the plate surface around immediately around the spine base is devoid of granulation. Both marginal plate (20–60 in number) series have rows of highly variable accessory granules present on the edges around them, similar to those on the abactinal plates. These granules range in shape from small and spiny, bead-like in outline, sharp and jagged to heavier and more blockish in shape ( Fig. 7C, E, I, J View Figure 7 ). As with the others, larger, blockier accessory granules occur closer together than those that are smaller, which are distributed further apart. A shallow but welldefined fasciolar groove occurs between marginals and is continuous from the abactinal surface to the actinal surface.
Marginals often have a large spine present on each plate (e.g. Fig. 7A, C, E, I View Figure 7 ). North Atlantic and North Pacific specimens tend to be lacking pedicelllariae on the marginal plate surface. However, other populations, such as from New Zealand, have one or two irregularly occurring bivalve pedicellariae with smoothedged valves, set into shallow pits on the plate surface ( Fig. 7E View Figure 7 ). Spines vary from one to three (both supero- and inferomarginal series), usually only one or two, and are similar in overall morphology to those present on the abactinal plates ( Fig. 7A, C, E, I View Figure 7 ). Spines vary in shape from blunt and cylindrical to sharp and conical. When more than one spine is present, the two or three spines emerge from a raised boss on the plate surface. Marginal plates with multiple spines tend to be found inter-radially. Sometimes, short tubercle-like spines sit adjacent or offset to spines. Marginal spines tend to be arranged in discrete, serial rows forming a spiny fringe around the marginal edge. Both marginal series but especially in inferomarginals, possess small, round granules present on marginal plate surface.
The actinal intermediate surface is broad and well developed, composed of six to eight actinal chevrons in an irregularly triangular pattern ( Fig. 7B View Figure 7 ). Plates round (proximally) to oval or irregular (distally adjacent to inferomarginal contact) in shape. Actinal plates covered by both large, prominent pedicellariae (e.g. Figs 7B View Figure 7 , 8B, J View Figure 8 ) and blunt to conical tubercles or spines with large accessory granules forming boundary around each plate. Size of accessory granules variable across individuals with larger blockier granules forming a mosaic on the actinal surface to spinelets, which form a prickly array on the oral surface. One to several spines or large, pointed granules present on actinal plates, sometimes occupying all plate surface. Some actinal plates bare with neither pedicellariae nor spine/ tubercle or large granule.
Actinal pedicellariae frequently bivalve, with smooth valves, large, occupying up to 85% of plate surface, sometimes bisecting the plate ( Fig. 7B View Figure 7 ). Abundance on actinal surface varies but it is common to see a full series of pedicellariae present on actinal plate series adjacent to the adambulacrals from the postoral to the edge of the disk with pedicellariae becoming more irregularly distributed more inter-radially, variably occupying anywhere from 40 to 90% of the actinal plates present on the actinal intermediate surface ( Fig. 7B View Figure 7 ).
Furrow spines blunt, thick to flattened and bladelike, numbering one or two, oval in cross-section but exceptionally three or four ( Fig. 7F View Figure 7 ). Three furrow spines sometimes present on postoral adambulacral plate. Subambulacral spine(s), thick, number one or two, sometimes replaced by pedicellariae. Adambulacral plate surface bare or covered with small to large granules, often blunt and round to angular and/or spinelet-like in appearance.
Oral plates with four to five blunt furrow spines, oval to flattened to triangular in cross-section. One large, blunt spine pointing into oral opening. Oral plate surface covered by large, blunt, cylindrical spines similar to those in furrow and subambulacral series in addition to smaller angular to rounded granules, six to nine adjacent to the actinal surface. Pedicellariae sometimes occurring in subambulacral positions behind furrow spines.
Widespread accounts indicate that H. phrygiana is a variably light orange to deep shade of orange or red (e.g. Fisher, 1911; Clark & Downey, 1992; Clark & McKnight, 2001) with white to yellow tube feet.
H. phrygiana from the Solomon Islands /Hawaii
Two specimens from the Central/South Pacific are new records. One from the SALOMONBOA 3 expedition ( IE- 2007-1311, Fig. 8 View Figure 8 A−C) is the first representative of Hippasteria collected from the Solomon Islands / Papua New Guinea region (722 m). DNA sampled from this specimen places it in the clade containing the widespread species, H. phrygiana , showing closest affinities with the Kerguelen Island, and to a lesser extent the New Zealand, population ( Foltz et al., 2013). A second specimen ( CASIZ 102006) from Hawaii at 396 m (1300 ft) shares several characters with and is very similar to MNHN IE- 2007-1311. However, CASIZ 102006 was fixed in formalin and unavailable for inclusion in our molecular data set.
Both specimens vary morphologically from North Atlantic/North Pacific/South Pacific H. phrygiana specimens in several respects. Abactinal, marginal, and actinal spines appear to be smaller and are more abun- dant, marginal plates have two rows of flattened granules forming perimeter around plate surface, pedicellariae are present on nearly every actinal plate, and the actinal pedicellariae themselves are composed of much narrower, more angular valves ( Fig. 8C View Figure 8 ). Actinal granules have more spinelet-like tips and are present on actinal plate surfaces as on the periphery ( Fig. 8B View Figure 8 ). In contrast, several characters are consistent with those of H. phrygiana , including the quadrateshaped marginal plates with only one to three (usually only one or two) spines on each plate, a single subambulacral spine, and only one to two (exceptionally three) furrow blunt, furrow spines. Both specimens also possess a large pedicellariae behind the subambulacrals on each adambulacral plate.
‘H. imperialis’ from Japan
Our data support a specimen ( CASIZ 115700) resembling H. imperialis Goto as part of the H. phrygiana clade. Morphology of this taxon shows some distinct differences from most members of H. phrygiana . The most notable distinctions come from the absence of the numerous conical spines on the abactinal surface ( Fig. 8H View Figure 8 ). ‘ Hippasteria imperialis ’ surface is covered by swollen, convex bosses or swellings with relatively few spines. Spines are present on the marginal plate surfaces of nearly all the other H. phrygiana populations, but are apparently absent from the marginal plate surface of H. imperialis ( Fig. 8H, I View Figure 8 ). Such a distinct difference in morphology suggests ecophenotypic variation, but further study is warranted.
Material examined
North Pacific. USNM E47392 View Materials H. aleutica paratype off north coast Kiska Island , Aleutians. 52°04′17″N, 177°15′20″E, 91.0 m. Coll. R GoogleMaps . Clark aboard FV (Fishing Vessel) Dominator, 31.vii.1997 (one dry spec. R = ∼5.2, r = 2.2); CASIZ 150478 ID as H. kurilensis Aleutian Islands , 52°22′46.56″N, 170°39′39.96″W to 52°22′1.1274″N, 170°39′2.52″W 278 m. Coll. M. Airey aboard F V GoogleMaps Vesteraalen 31.v.2000, NMFS Aleutian Survey 2000 (one dry spec. R = 6.3, r = 3.5); CASIZ 141112 ID as H. spinosa Aleutian Islands 52°4′32.0514″N, 176°21′17.6394″W to 52°4′55.452″N, 176°20′15.36″W, 149 m. Coll. M. Airey aboard F V GoogleMaps Vesteraalen 31.v.2000, NMFS Aleutian Survey 2000 (one dry spec. R = 4.9, 4 = 3.5, arms upturned); CASIZ 117923 ID as H. spinosa . Aleutian Islands 52°54.9624′N, 169°52.056′W, 106 m, coll. B. Frye aboard F V GoogleMaps Vesteraalen , 16.v.1999 (one dry spec. R = 8.2, r = 4.5); CASIZ 101829 ID as H. spinosa off Shelter Cove, Humboldt County, CA 39°53′N, 124°07′W, 183 m (100 fms). Coll. R GoogleMaps . Bolin , 5.ix.1935 (one dry spec. R = 6.5, r = 4.9); CASIZ 101830 as H. spinosa off Shelter Cove, Humboldt County, CA 39°53′N, 124°07′W, 183 m (100 fms). Coll. R GoogleMaps . Bolin , 5.ix.1935 (one dry spec. R = 10.6, r = 6.0); CASIZ 107744 . Id as H. spinosa Hein Bank, Strait of Juan de Fuca , Puget Sound, Washington, 48°19′N, 123°5′W, 45.7 m (25 fms). Coll. E. Swan, 6.viii.1951 (one dry spec. R = 15.6. r = 7.3); USNM 1215324 About USNM GoogleMaps President Jackson Seamount C, North Pacific. 42°44′50.4″N, 128°5′25.5″W (42.747347, −128.09044), 1405.8 m, coll. MBARI D82 - A21 (one wet spec. R = 3.9, r = 2.1) GoogleMaps ; CASIZ 115674 Id as H. spinosa . San Pedro Channel , off Avalon , Santa Catalina Island, Channel Islands 33°23′N, 118°20′W, 146 m (80 fms). Coll. Myers et al. on the Zaca, 17.ix.1938 (one wet spec. R = 4.0, r = 2.5); CASIZ 115695 GoogleMaps Id as H. spinosa . Off Santa Catalina Island, Channel Islands 33°22′N 118°20′W, 60 m. Coll. 20.ix.1938 (one wet spec. R = 5.2, r = 3.3); CASIZ 102006 GoogleMaps 6 miles offshore from Makapuu Point. Oahu , Hawaii. 396 m (1300 ft). Coll. S. Earle, B. Bartko fr. Deep Star II. 25.x.1979 (one dry spec. R = 12.5, r = 7.6) .
South Pacific. Hippasteria hyadesi holotype. EcAs 2840 south-east of Port Famine (Puerto del Hambre), Chile, 100 m (one dry spec. R = 4.7, r = 2.2, specimen is partly damaged); Id as H. hyadesi USNM E 13586 northwest of Amundsen Sea, South Pacific. 54°49′S, 129°48′W, 549 m. Coll. R V Eltanin (three dry specs R = 3.5, r = 2.0; R = 8.2, r = 3.9; R = ∼9.4, r = 4.7, arms upturned). Id as H. trojana USNM E 13768 east of Banks peninsula, South Island, New Zealand. 43°54′S 174°35′54″E, 585 m. Coll. 27.viii.1971 (one dry spec. R = ∼10.1, r = 6.1, arms upturned); Id as H. hyadesi USNM 1082740 South Pacific (sub-Antarctic) 67°23′S− 67°24′S to 180°00′W – 179°58′W, 595– 516 m. Coll. US Antarctic Research Program (two dry specs R = 8.3, r = 4.3; R = 10.1, r = 6.2); USNM 1121154 Cape Froward, Strait of Magellan. 53°39′24″S, 70°55′W, 82 m (one dry spec. R = 8.4, r = 4.5). MNHN IE- 2007-1311 Solomon Islands 9°12′S, 160°55′E, 722 m, coll. Richer de Forges & Boisselier, SALOMONBOA 3 aboard N/O Alis (one wet spec. R = 12.4, r = 7.6).
South Indian Ocean. EcAs 10692. Id as H. hyadesi . Crozet Islands 45°37′S, 50°34′E, 200 m. Coll. N.O. Marion Dufresne MD 30 GoogleMaps , 20.ii.1982 (six dry specs R = 11.4, r = 6.2; R = 8.6, r = 4.6; R = 8.2, r = 4.3; R = 6.7, r = 3.8; R = 6.6, r = 3.3; R = 6.9, r = 3.3); EcAs 10683. Id as H. hyadesi Crozet Islands 46°33′S, 51°47′E, 200 m. Coll. N.O. Marion Dufresne MD 30 GoogleMaps , 24.ii.1982 (five dry specs R = 4.6, r = 2.6; R = 3.8, r = 1.8; R = 2.8, r = 1.5; R = 3.0, r = 1.4; R = 2.8, r = 1.6); EcAs 10860. Id as H. hyadesi . Crozet Islands, 210 m. Coll. N.O. Marion Dufresne MD 08 , CP 257 (one dry spec. 13.1, r = 7.7); EcAs 10861. Crozet Islands, 46°26′S, 52°04′E, 50 m. Coll. N.O. Marion Dufresne MD 08 GoogleMaps , CP DC 309 (two dry specs R = 7.7, r = 3.6; R = 16.1, r = 7.0); MNHN IE- 2006-1162 Id as H. hyadesi . Kerguelen Islands 48°43′S, 71°06′E, 925– 937 m. Coll. A. Guille, 13.iii.1975 (one dry spec. R = 10.5, r = 6.5); MNHN IE-2006-1154 GoogleMaps Id as H. hyadesi . Kerguelen Islands 52°43.26′S, 72°41.73′E, 240 m. Coll. N.O. Marion Dufresne 42, st. 02–12 (one dry spec. R = 13.1, r = 8.2), EcAs 10735. Marion Island. 46°41′S, 38°73′E, 315– 570 m. Coll. N.O. Marion Dufresne MD08 GoogleMaps , CP173 29.iii.1976 (one dry spec. R = 8.7, r = 3.9); EcAs 10856 Marion Island. 46°47′S, 38°03′E, 180 m. Coll. N. O. Marion Dufresne MD08 GoogleMaps , cp116 26.iii.1976 (one dry spec. R = 8.9, r = 4.7), EcAs 10858 Marion Island. 46°45′S, 37°57′E, 185– 232 m. Coll. N. O. Marion Dufresne MD08 GoogleMaps , CP134 27.iii.1976 (one dry spec. R = 11.2, r = 7.1); EcAs 10854 Bouvet Island 53°13′S, 44°39′E, 570– 585 m. Coll. N. O. Marion Dufresne MD24 GoogleMaps , CP51 2.ix.1980 (one dry spec. R = 6.4, r = 2.8), EcAs 10852 Bouvet Island 46°1′S 42°32′E, 270– 315 m. Coll. N. O. Marion Dufresne MD24 , CM69 GoogleMaps 8.ix.1980 (three dry specs R = 6.7, r = 3.4, R = 7.4, r = 3.2; R = 1.5, r = 1.0) .
South Atlantic /Sub-Antarctic. Id as H. hyadesi USNM E 43921 66°20′S, 67°47′W, 325 m. Coll. 25.iii.1959 (one dry spec. R = 7.4, r = 3.5) GoogleMaps .
North Atlantic. USNM 14361 About USNM Georges Bank, Massachusetts , 287 m (157 fms), coll. USFC Albatross , 30.viii.1883 (one dry spec. R = 10.8, r = 6.1); USNM E46615 View Materials northern edge of Georges Bank. 42°7′N, 66°23′W, 82 m. coll. R V GoogleMaps Albatross IV (one dry spec. R = 6.3, r = 3.3); USNM E46606 View Materials , south-east of Cape Elizabeth, Gulf of Maine, Maine. 43°31′N, 69°49′W, 97 m. Coll. R V GoogleMaps Albatross IV, 14.vii.1965 (one dry spec. R = 6.2, r = 3.7); USNM E 46610 View Materials , Browns Bank , Nova Scotia, 42°34′N, 65°44′W, 93 m, coll. R V GoogleMaps Albatross IV, 15.x.1965 (one dry spec. R = 7.3, r = 3.5); USNM E 46611 View Materials Browns Bank , Nova Scotia, Canada. 42°45′N, 65°7′W. 97.0 m. Coll. R V GoogleMaps Albatross IV, 15.x.1965 (one dry spec. R = 1.7, r = 1.0). USNM E46613 View Materials western part of Georges Bank, Massachusetts. 41°24′N, 68°25′W, 68 m. Coll. R V GoogleMaps Albatross IV, 28.x.1965 (one dry spec. R = 2.2, r = 1.7); USNM E46619 View Materials south of Browns Bank , Nova Scotia 42°11′N, 65°50′W, 247 m. Coll. R V GoogleMaps Albatross IV (12 dry specs R = 5.1, r = 2.4; R = 4.4, r = 2.2; R = 5.1, r = 2.8; R = 4.3, r = 2.0; R = 3.3, r = 1.6; R = 3.2, r = 1.6; R = 3.0, r = 1.5; R = 3.0, r = 1.5; R = 3.1, r = 1.2; R = 2.9, r = 1.5; R = 2.7, r = 1.4; R = 2.2, r = 1.2); USNM 46622 About USNM south of Browns Bank , Nova Scotia 42°2′N, 65°47′W, 256 m. Coll. R V GoogleMaps Albatross IV (two dry specs R = 6.2, r = 2.7; R = 4.2, r = 2.0). USNM E30719 View Materials Baltimore Canyon , 38°9′30″N, 73°51′12″W, 457 m, coll. Bureau of Land Management/ Johnson Sea Link. iii.1983 (one dry spec. R = 15.5, r = 8.3); USNM E30720 View Materials GoogleMaps Baltimore Canyon , 38°9′56″N, 73°51′52″W, 417 m, coll. Bureau of Land Management/ Johnson Sea Link, iii.1983 (one dry spec. R = ∼13.7, r = 7.2) GoogleMaps .
Alton M. 1966. Bathymeric distribution of sea stars (Asteroidea) off the Northern Oregon coast. Journal of the Fisheries Research Board of Canada 23: 1673 - 1714.
Anisimova NA, Cochrane SJ. 2003. An annotated checklist of the echinoderms of the Svalbard and Franz Josef Land archipelagos and adjacent waters. Sarsia 88: 113 - 135.
Baranova ZI. 1957. Echinoderms of the Bering Sea. Issledovaniya Dalny-Vostok Morei USSR 4: 149 - 266 (in Russian).
Barrett L. 1857. Descriptions of four new species of Echinodermata. Annals of the Magazine of Natural History 20: 46 - 48.
Bell FJ. 1892. Catalogue of the British Echinoderms in the British Museum (Natural History). London, 16 pls.
Bernasconi I. 1961. Una nueva especie de asteroideo. Neotropica 7: 1 - 2.
Bernasconi I. 1963. Asteroideos Argentinos. IV. Familia Goniasteridae. Revista del Museo Argentino de Ciencias naturales (Zoologia) 9: 58 - 89.
Blacker RW. 1957. Benthic animals as indicators of hydrographic conditions and climate change in Svalbard waters. Fishery Investigations. Ministry of Agriculture, London 20: 1 - 49.
Branch ML, Jangoux M, Alva V, Massin CL, Stampanato S. 1993. The Echinodermata of subantarctic Marion and Prince Edward Islands. South African Journal of Antarctic Research 23 (1 & 2): 37 - 70.
Buchanan JB. 1966. The marine fauna of the Cullercoats district. D. Echinodermata. Report of the Dove Marine Laboratory 3: 21 - 39.
Carey AG. 1972. Food sources of sublittoral, bathyal and abyssal asteroids in the Northeast Pacific Ocean. Ophelia 10: 35 - 47.
Clark HL. 1905. Fauna of New England. 4. List of the Echinodermata. Occasional Papers of the Boston Society of Natural History 7: 1 - 16.
Clark HL. 1923. The Echinoderm fauna of South Africa. Annals of the South African Museum 13: 221 - 435.
Clark HL. 1926. Echinoderms from the South African Fisheries and Marine Biological Survey. 2. Sea-stars. Reports of the Fisheries and Marine Biological Survey of the Union of South Africa 4, sp. report 7: 1 - 33.
Clark AH. 1949. Echinoderms from the mid-Atlantic dredged by the Atlantis in the summer of 1948. Journal of the Washington Academy of Science 39: 371 - 377.
Clark AM. 1952. Some echinoderms from South Africa. Transactions of the Royal Society of South Africa 33: 193 - 221.
Clark AM. 1962. Asteroidea. B. A. N. Z. Antarctic Research Expedition 1929 - 1931 B 9: 1 - 104.
Clark HES. 1970. Sea-stars (Echinodermata: Asteroidea) from Eltanin cruise 26, with a review of the New Zealand asteroid fauna. Zoology Publications from Victoria University of Wellington 52: 1 - 33.
Clark AM, Courtman-Stock J. 1976. The echinoderms of Southern Africa. London: British Museum of Natural History.
Clark AM, Downey ME. 1992. Starfishes of the Atlantic. London: Chapman and Hall.
Clark AM. 1993. An index of names of recent Asteroidea - part 2: Valvatida. Echinoderm Studies 4: 187 - 366.
Clark HES, McKnight DG. 2001. The marine fauna of New Zealand: Echinodermata: Asteroidea (sea-stars), order Valvatida. NIWA Biodiversity Memoir 117: 1 - 270.
Clark RN, Jewett SC. 2011. A new sea star of the genus Hippasteria (Asteroidea: Goniasteridae) from the Aleutian Islands. Zootaxa 2963: 48 - 54.
Codoceo MR, Andrade H. 1978. Asterozoos arquibentonicos de Chile Central. Anales del Museo de Historia Natural de Valparaiso 11: 153 - 174.
Danielssen DC, Koren J. 1882. Fra den norske Nordhavsexpedition: Echinodermer. Nyt Magazin for Naturvidenskaberne 26: 267 - 299.
Dilman A. 2006. Asteroid fauna of the Reykjanes Ridge. In: Mironov AN, Gebruk AV, Southward AJ, eds. Biogeography of the North Atlantic seamounts. Moscow: KMK Scientific Press Ltd., 177 - 192, 196 pp.
Djakonov AM. 1950. Morskie Zvezdy Morei SSSR Tableaux analytiques de la faune de U. R. S. S. 34: 1 - 203. [Translated as Dyakonov, A. M. 1968. Sea stars (Asteroids) of the USSR Seas. Keys to the Fauna of the USSR 34. Zoological Institute of the Academy of Sciences of the USSR, A. A. Strelkov. ed. Israel Program for Scientific Translations Ltd. Jerusalem].
Doderlein L. 1900. Die Echinodermen. Zoologische Ergebnisse einer Untersuchungsfahrt des deutschen Seefischerei- Vereins nach der Bareninsel und Westspitzbergen ausgefuhrt im Sommer 1898 auf S. M. S Olga. Helgolander Wissenschaft Meeresunters 4: 195 - 248, pls. 4 - 10.
Dons C. 1937. Zoologische Notizen XXXIV. Hippasteria insignis n. sp. Det Kongelige Norske Videnskabers selskab Forhandlinger 10: 16 - 19, 161 - 164; 165 - 168.
Duben MW, Koren J. 1846. Ofversigt af Skandinaviens Echinodermer. Ofversigt af K. Vetenskapsakademiens forhandlingar 1845: 73 - 77.
Einarsson H. 1948. Echinoderma. The Zoology of Iceland 4: 1 - 67.
Fell HB. 1958. Deep-sea echinoderms of New Zealand. Zoology Publications from Victoria University of Wellington 24: 1 - 40.
Fisher WK. 1911. Asteroidea of the North Pacific and adjacent waters. 1. Phanerozonia and Spinulosida. Bulletin of the US National Museum 76: xiii + 420 pp, 122 pls.
Foltz DW, Fatland SD, Eleaume M, Markello K, Howell KL, Neill K, Mah CL. 2013. Global population divergence of the sea star Hippasteria phrygiana corresponds to the onset of the last glacial period of the Pleistocene. Marine Biology 160: 1285 - 1296.
Forbes E. 1841. A history of British starfishes and other animals of the class Echinodermata. London: John Van Voorst.
Forbes E. 1843. On a new British starfish of the genus Goniaster (G. abbensis). Annals of the Magazine of Natural History 11: 280 - 281.
Franz DR, Worley EK, Merrill AS. 1981. Distribution patterns of common seastars of the Middle Atlantic continental shelf of the Northwest Atlantic (Gulf of Maine to Cape Hatteras). Biological Bulletin 160: 394 - 418.
Fujikura K, Okutani T, Maruyama T. 2008. Deep-sea life-Biological Observations Using Research Submersibles. Tokai University Press. 487 pp. (ISBN 978 - 4 - 486 - 01787 - 5 in Japanese).
Ganong WF. 1893. Zoological notes. Report of the Committee on Marine Invertebrate Zoology. Bulletin of the Natural History Society of New Brunswick 9: 46 - 59.
Gish RK. 2007. The 2006 Petrel Bank red king crab survey. Fishery Management Report (Alaska) 07 - 44: 1 - 90.
Goto H. 1914. A descriptive monograph of Japanese Asteroidea. 1. Journal of the College of Sciences, Imperial University of Tokyo 29: 1 - 808.
Gray JE. 1840. XXXII. A Synopsis of the genera and species of the Class Hypostoma (Asterias, Linnaeus). Annals and Magazine of Natural History 1: 275 - 290.
Gray JE. 1866. Synopsis of the species of starfish in the British Museum. Iv. + 17 London.
Grieg JA. 1895. On echinodermen-faunaen I de Vestlandiske Fjorde. Bergens Museums Aarbok 1894 - 95: 1 - 13.
Grieg JA. 1902. Oversigt over det nordlige Norges Echinodermer. Bergens Museums Aarbok 1902: 1 - 38, 1 pl., 1 fig.
Grieg JA. 1907. Echinodermen von dem norwegischen Fischereidampfer Michael Sars in den Jahren 1900 - 1903 gesammelt. 3. Asteroidea. 1. Bergens Museums Aarbok 1906: 1 - 87, 2 pls.
Grieg JA. 1912. Sognefjordens echinodermer. Archiv foer Matematik og Naturvidenskab 32: 1 - 13.
Grieg JA. 1913. Bidrag til kundskapen om Hardangerfjordens fauna. Bergens Museums Aarbok 1913: 1 - 147, 2 pls. (Echinodermata, 108 - 140).
Grieg JA. 1917. Echinodermen indsamlet av Michael Sars sommeren 1914. Archiv foer Matematik og Naturvidenskab 34: 1 - 11.
Grieg JA. 1921. Echinodermata. Report on the Scientific Results of the ' Michael Sars' North Atlantic Expedition, 1910. Bergen (1921) 3 (2): 1 - 47, 5 pls, 10 figs.
Gulliksen B, Palerud R, Brattegard T, Sneli JA. 1999. Distribution of marine benthic macro-organisms at Svalbard (including Bear Island) and Jan Mayen. Figures and tabulated catalogues of species-distributions, literature and synonyms. Research Report for DN 1999 - 4. Trondheim, Norway: Directorate for Nature Management.
Hartlaub C. 1900. Zoologische Ergebnisse einer Untersuchungsfahrt des deutschen Seefischerei Vereins nach der Bareninsel und Westspitzbergen ausgefuhrt im Sommer 1898 auf S. M. S. Olga. 1. Helgolander Wissenschaft Meeresunters 4: 171 - 193.
Haubold S. 1933. Uber eine neue Form sitzender Pedizellarien bei Seestern. Zoologischer Anzeiger 103: 199 - 205.
Hayashi R. 1952. Sea-stars of Seto and adjacent waters. Publications of the Seto Marine Biological Laboratory 11: 143 - 159.
Imaoka T, Irimura S, Okutani T, Oguro C, Oji T, Shigei M, Horikawa H. 1990. Echinoderms from the continental shelf and slope around Japan, Vol. I. Tokyo: Japan Fisheries Resource Conservation Association.
Johnston G. 1836. Illustrations in British Zoology: Asterias rubens and A. johnstoni Gray; Asterias aranciaca and A. endeca; Asterias papposa. Magazine of Natural History 9: 144 - 147, 298 - 300, 474 - 475.
Koehler R. 1909. An account of the deep-sea Asteroidea collected by the Royal Indian Marine Survey ship Investigator. Echinoderma of the Indian Museum Part 5: 115 - 131.
Koehler R. 1924. Les Echinodermes des Mers d'Europe. 1. Generalites, Asterides, Ophiurides. Paris: Librairie Octave Doin, xiii. + 360 pp, 9 pls.
Koehler R. 1926. Echinodermata; Echinoidea. Australasian Antarctic Expedition 1911 - 1914. Scientific reports. Series C, zoology and botany. Sydney: David Harold Paisley, Government Printer, 8 (3): 1 - 134.
Koslow JA, Gowlett-Holmes K. 1998. The seamount fauna off southern Tasmania: benthic communities, their conservation and impacts of trawling. Final report to Environment Australia & the Fisheries Research Development Corporation. Hobart: CSIRO.
Lamarck JBPA de. 1816. Stellerides. Histoire naturelle des animaux sans vertebres, 1 st edn. Paris. 2: 522 - 568
Lambert P. 1978 a. British Columbia Marine Faunistic Survey Report: asteroids from the Northeast Pacific. Fisheries and Marine Service Technical Report 773: 1 - 23.
Lambert P. 2000. Sea stars of British Columbia, Southeast Alaska, and Puget Sound. Royal British Columbia Museum Handbook. Vancouver: University of British Columbia Press.
Larrain A, Mutschke E, Riveros A, Solar E. 1999. Preliminary report on Echinoidea and Asteroidea (Echinodermata) of the joint Chilean-German-Italian Magellan Victor Hensen Campaign, 17 Oct- 27 November 1994. Scientia Marina 63 (Suppl. 1): 433 - 438.
Linck JH. 1733. De Stellis Marinis liber singularis. xxiv + 107 pp. Lipsiae.
Ludwig H. 1900. Arktische Seesterne. Fauna Arctica 1: 447 - 502.
Mah C. 1998 a. New records, taxonomic notes and a checklist of Hawaiian starfish. Bishop Museum Occasional Papers 55: 65 - 71.
Mah CL, Nizinski M, Lundsten L. 2010. Phylogenetic revision of the Hippasterinae (Goniasteridae; Asteroidea): systematics of deep sea corallivores, including one new genus and three new species. Zoological Journal of the Linnean Society 160: 266 - 301, 11 figs.
Mah C, Hansson H. 2012. Hippasteria. In: Mah CL, ed. World Asteroidea database. Accessed through: Mah CL. (2012). World Asteroidea database at http: // www. marinespecies. org / Asteroidea / aphia. php? p = taxdetails & id = 123297 (last accessed on 16 July 2012).
Maluf Y. 1988. Composition and distribution of the central eastern Pacific echinoderms. Natural History Museum of Los Angeles County, Technical Reports 2, Los Angeles, California.
McKnight DG. 1967. Additions to the echinoderm fauna of the Chatham Rise. New Zealand Journal of Marine and Freshwater Resources 1: 291 - 313.
Moore JA, Vecchione M, Collette BB, Gibbons R, Hartel KE. 2004. Selected fauna of Bear Seamount (New England Seamount chain), and the presence of ' natural invader' species. Archive of Fishery and Marine Research 51: 241 - 250.
Mortensen T. 1927. Handbook of the echinoderms of the British Isles. London: Oxford University Press.
Mortensen T. 1932. Echinooderms of the Godthaab Expedition of 1928. Meddelelser om GrOnland 79: 1 - 62.
Mortensen T. 1933. Echinoderms of South Africa (Asteroidea and Ophiuroidea). Videnskabelige Meddelelser fra Dansk naturhistorisk Forening i. Kobenhavn 93: 215 - 400.
Muller J, Troschel FH. 1842. System der Asteriden. 1. Asteriae. 2. Ophiuridae. Braunschweig, 12 pls.
Nordgaard O. 1905. Hydrographical and biological investigations in Norwegian fiords. 3. Bottom life. Bergens Museums Skrifter 7: 153 - 256, 21 pls, 10 figs.
Norman AM. 1865. On the genera and species of British Echinodermata. 1. Crinoidea, Ophiuroidea, Asteroidea. Annals of the Magazine of Natural History 15: 98 - 129.
O'Connor B, Tyndall P. 1986. Notes on eight species of rare or deep water echinoderms from Irish inshore waters. Irish Naturalists Journal 22: 96 - 97.
Parelius J. 1768. Beskrivelse over Nogle Korstrold. Kongelige Norske Videnskabers Selskab Skrifter 4: 423 - 428, pl. 14.
Pearcey FG. 1902. Notes on the marine deposits of the Firth of Forth, and their relation to its animal life. Transactions of the Society of Glasgow (n. s.) 6: 217 - 251.
Perrier E. 1875. Revision de la collection de stellerides du Museum d'Histoire naturelle de Paris. Paris: Reinwald.
Perrier E. 1888. Sur la collection d'etoiles de mer recueillie par la Commission Scientifique du Cap Horn. Comptes Rendus Hebdomadaires des Seances de l'Academie des Sciences. Paris 106: 763 - 765.
Perrier E. 1891. Echinoderma I. Stellerides. Mission Scientifique du Cap Horn: 1882 - 83. 6. Zoologie (3), Paris.
Picton BE. 1993. A field guide to the shallow-water echinoderms of the British Isles. London: Immel Publishing.
Rowe FWE, Gates J. 1995. Echinodermata. In: Wells A, ed. Zoological catalogue of Australia 33. Melbourne, Australia: CSIRO, xii: 1 - 510.
Simpson J. 1903. Catalogue of echinoderms found at Aberdeen and neighbourhood. Transactions of the Aberdeen Working Mens Natural History Society 1: 39 - 43.
Sladen WP. 1883. Asteroidea dredged in the Faeroe Channel during the cruise of H. M. S. Triton in August, 1882. Transactions of the Royal Society of Edinburgh 32: 153 - 164.
Sladen WP. 1889. Asteroidea. Report of the Scientific Results of H. M. S. Challenger 30: 1 - 893.
Stampanato S, Jangoux M. 2004. The asteroid fauna (Echinodermata) of Marion and Prince Edward Islands. Annals of the South African Museum 112: 1 - 16.
Sussbach S, Breckner A. 1911. Die Seeigel, Seesterne und Schlangensterne der Nord- und Ostsee. Helgolander Wissenschaft Meereesunters (n. s.) 12: 167 - 300, 3 pl.
Verrill AE. 1874. Report upon the invertebrate animals of Vineyard Sound, etc., Washington 1873. Report of the Commissioner of Fish and Fisheries for 1871. 295 - 778, pls I-XXXVIII, 1873 - 74.
Verrill AE. 1895. Distribution of the echinoderms of north-eastern America. American Journal of Science 49: 127 - 141.
Verrill AE. 1899. Revision of certain genera and species of starfishes, with descriptions of new forms. Transactions of the Connecticut Academy of Arts and Sciences 10: 145 - 234.
Verrill AE. 1909. Descriptions of new genera and species of starfishes from the North Pacific coast of America. American Journal of Science 28: 59 - 70.
Verrill AE. 1914. Monograph of the shallow-water starfishes of the North Pacific coast from the Arctic Ocean to California. Harriman Alaska Series: US National Museum 14: 1 - 408.
Walker CW. 1978. Studies on the reproductive systems of seastars. 3. Preliminary report on the morphology, histology and ultrastructure of the gonad and gonoduct of the horse-star Hippasteria phrygiana (Asteroidea, Goniasteridae). Thalassia Jugoslavica 12: 361 - 369.
Whiteaves JF. 1901. Catalogue of the marine Invertebrata of Eastern Canada. Ottawa: Geological Survey of Canada, 1 fig.
Wolff WJ. 1968. The Echinodermata of the estuarine region of the rivers Rhine, Meuse and Scheldt with a list of species occurring in the coastal waters of the Netherlands. Netherlands Journal of Sea Research 4: 59 - 85.

Figure 1. Evolutionary relationships of sampled Hippasterinae (Goniasteridae) based on 654 aligned nucleotide positions of the cytochrome c oxidase subunit I gene. The phylogenetic tree was inferred using the neighbour-joining method (Saitou & Nei, 1987) in MEGA v. 5.2.1 (Tamura et al., 2011). The bootstrap consensus tree inferred from 500 replicates was drawn to scale, with distances computed using the maximum composite likelihood method (Tamura et al., 2004), in units of numbers of base substitutions per site. Ambiguous positions were removed for each pair of sequences prior to calculating the distance matrix. Terminals were labelled with GenBank accession numbers and (where available) with museum voucher numbers. Bootstrap support values are shown above or below branches when greater than 70%.
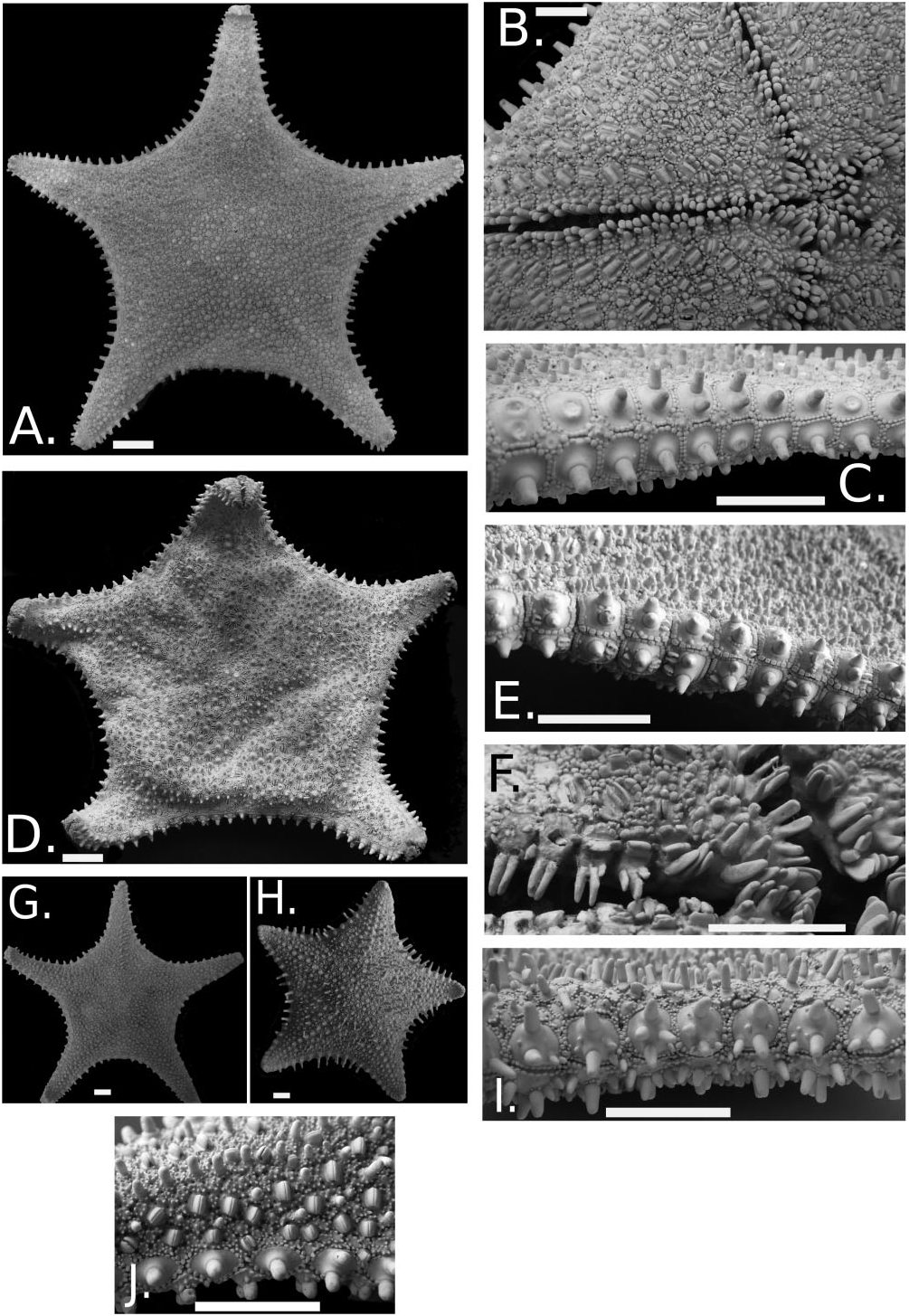
Figure 7. Hippasteria phrygiana (North Atlantic, South Pacific). USNM 14361 North Atlantic. Scale bar = 1.0 cm. A, abactinal surface. B, actinal surface. C, marginal plate series. USNM E13768 New Zealand. D, abactinal surface. E, marginal plate series. F, actinal surface showing furrow and mouth spines. G and H show USNM E46619 and MNHN (Paris) EcAs 10683, respectively, at R = ∼4.5–5.0. I, CASIZ 113374 British Columbia. J, CASIZ 173038 Aleutians. Lateral view showing abactinal spines, pedicellariae, and marginal plates.
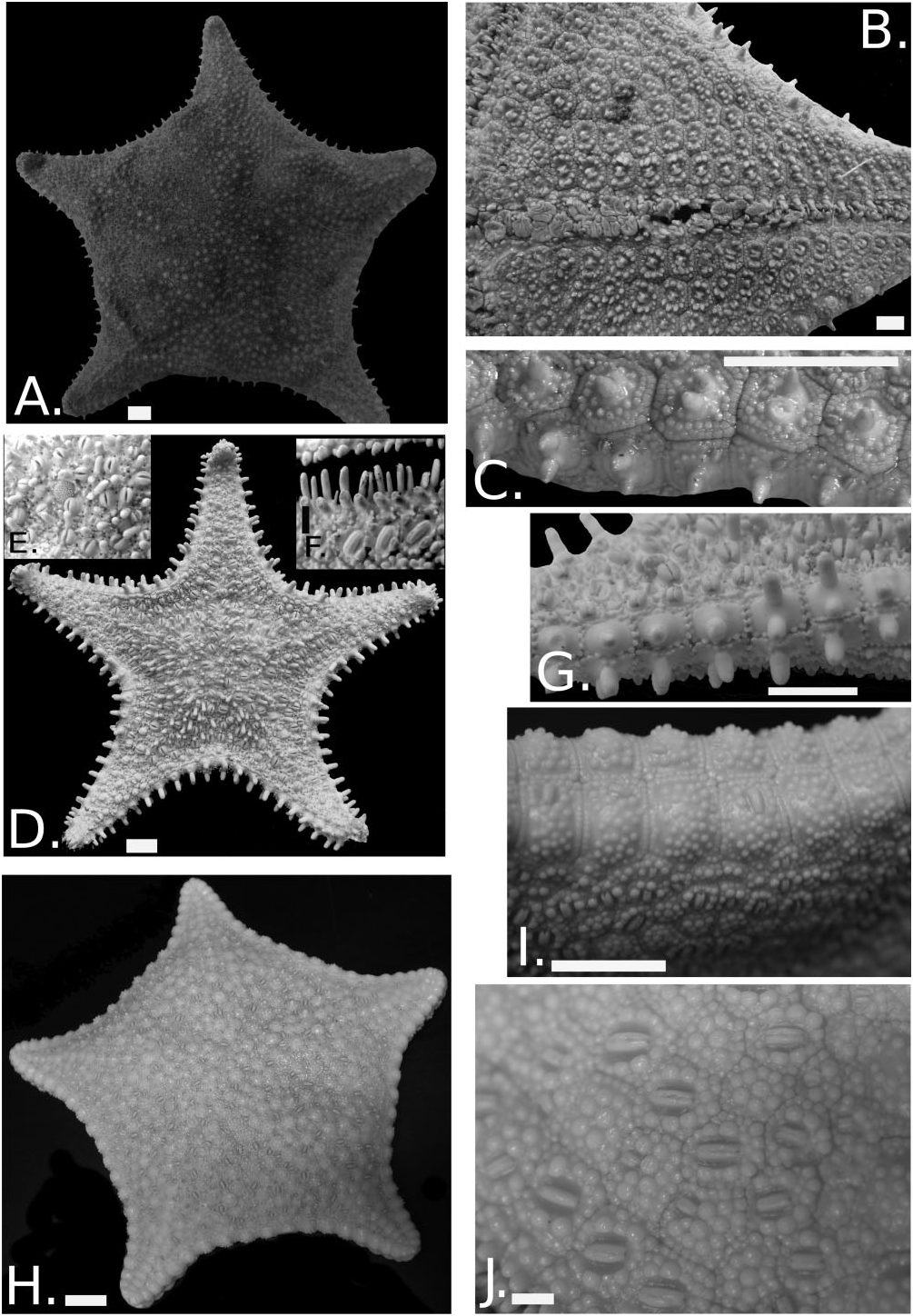
Figure 8. Hippasteria phrygiana variation across its range. Sclae bar = 1.0 cm for all unless specified otherwise. A, MNHN IE-2007-1311. Solomon Islands. Abactinal surface. B, actinal surface. C, marginal plates. D, Hippasteria aleutica paratype USNM E47392. E, abactinal spines and pedicellariae. Each pedicellariae is 2.0 mm in length. F, furrow spines and actinal pedicellariae. Scale bar = 0.5 cm. G, marginal plates. H, CASIZ 115700 Hippasteria imperialis. Abactinal surface. I, marginal plates. J, actinal surface showing pedicellariae. Scale bar = 0.5 cm.
| USNM |
Smithsonian Institution, National Museum of Natural History |
| NOAA |
National Oceanic and Atmospeheric Administration |
| LACM |
Natural History Museum of Los Angeles County |
| R |
Departamento de Geologia, Universidad de Chile |
| MNHN |
Museum National d'Histoire Naturelle |
| MCZ |
Museum of Comparative Zoology |
| IE |
Cepario de Hongos del Instituto de Ecologia |
| NIWA |
National Institute of Water and Atmospheric Research |
| V |
Royal British Columbia Museum - Herbarium |
| CA |
Chicago Academy of Sciences |
| US |
University of Stellenbosch |
| USFC |
U. S. Fish Commission |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hippasteria
| Mah, Christopher, Neill, Kate, Eléaume, Marc & Foltz, David 2014 |
Hippasteria nozawai
| Goto 1914 |
H. spinosa
| Verrill 1909 |
HIPPASTERIA PHRYGIANA ( PARELIUS, 1768 )
| PHRYGIANA (PARELIUS 1768 |
Asterias phrygiana
| Parelius 1768 |
Hippasteria
| PHRYGIANA (PARELIUS 1768 |