
Simrothiella vasconiensis, Salvini-Plawen, 2008
|
publication ID |
https://doi.org/10.5281/zenodo.4525775 |
|
persistent identifier |
https://treatment.plazi.org/id/0394ED6F-FFA8-0118-EC34-FBD2248DFD2C |
|
treatment provided by |
Felipe (2021-02-09 14:07:20, last updated by Plazi 2023-11-29 20:37:53) |
|
scientific name |
Simrothiella vasconiensis |
| status |
sp. nov. |
Simrothiella vasconiensis View in CoL n. sp.
( Figs 1-9 View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG View FIG )
HOLOTYPE. — Sclerites, section series on slides; midbody 2 × in araldite ( MNHN 9833 About MNHN ).
PARATYPES. — 1) Cap Breton 88, stn DE 07, section series on slides, midbody 2 × in araldite ( MNCN 15.02 About MNCN /0017) ; 2) Cap Breton 88, stn DE 07, section series on slides; midbody in araldite ( MNHN 9834 About MNHN ) .
TYPE LOCALITY. — Southwestern France, Atlantic coast, Cap Breton 88, stn DE 05.
ETYMOLOGY. — Referring to the ancient name Vasconia for the Basque area bordering the southeastern golfe de Gascogne (Bay of Biscay).
OTHER MATERIAL EXAMINED. — France. Cap Breton 88, stn DE 05, 43°57.42’N, 02°05.16’W, 164 m, 5.VII.1988, Jean-Claude Sorbe and Serge Gofas, 11 specimens (incl. 6 larger fragments), mostly rolled up (MNHN). — Same data, stn DE 07, 43°58.65’N, 02°05.27’W, 170 m, 8 specimens (incl. a fragment).
The body ends of two specimens from each station were serially cross-sectioned (cs 1-1.5 µm) in araldite and stained with toluidine blue. One fragment from DE 05 was manually dissected for preparation of the radula apparatus by histolysing tissues with bleach (hypochlorite solution).
Material examined for comparison: Simrothiella margaritacea (Koren & Danielssen, 1977) of Odhner’s (1921) section series (SMNH 4731) was examined: 4731:1 (half anterior end, cs 5 µm), 4731:2a-c (cs 10 µm), 4731: 3a-c (posterior body, cs 5 µm) and 4731:4a, b (ls 5 µm and 10 µm); the series 4731:1 and 4731:3 are not complete, but series 4731:2 and 4731:4 should be regarded as part of the type material of this species (cf. Salvini-Plawen 2004).
DIAGNOSIS. — Up to 7.5 mm long. Mantle with epidermis papillae. Single pedal fold does not enter mantle cavity; a strong pallial hook at both sides of anterior cavity opening; mantle cavity with paired ventro-anterior pouch or sack, respiratory organs papilla-like or missing. Radula plates
with 15 heterogeneous distal denticles; radula support (bolster, odontophore) with turgescent vesicles; midgut caecum frontally paired. Supra-rectal commissure short. Spawning ducts with a blind central portion before their junction. At each side a long copulatory stylet and a short, more scaly element.
DESCRIPTION
External appearance
Up to 7.5 mm long, fairly tubular, but anterior part of the body somewhat wider, posterior part may be club-shaped; in preserved state often rolled up ( Fig. 1A View FIG ). Sclerites almost adjacent but causing a slightly uneven outline. Closer inspection reveals multilayered, intercrossing spicules; body openings inconspicuous, pedal groove visible.
Mantle
Epithelium 6-12 µm thick; with several slender papillae, distal “head” up to 10 µm diam.; cuticle 30 µm (ventrally) to 50 µm thick. Sclerites consisting of acicular hollow needles, slightly curved or straight or bent, 70-170 × 7-14 µm ( Fig. 2 View FIG ); they are thickwalled (2.5-3.5 µm) in cross section, yielding only a 4-7 µm wide cavity, and intercross in two or three layers.Ventrally, lateral to the pedal groove, the hollow needles measure only 40 × 5 µm; adjacent to the foot, there are knife-blade scales (45-60 × 10 µm).
Foot and mantle cavity
Pedal pit shallow, posteriorly bilobed; middorsal roof continues into one low pedal fold, which ends anterior to the opening of the mantle cavity proper, separated by a small pit with folded epithelium (not continuous with the pedal fold). Pedal gland consists of two different glandular cell types densely grouped together: first, pale-stained cells of reticulate appearance, which open frontally and laterally into pedal pit; second, cells with dense, deeply stained granula opening dorsally into pit. The latter type continues in paired arrangement as smaller sole glands along pedal groove, where they open intercellularly within the pedal fold.
Mantle cavity opening entirely ventral, epithelium in posterior half of mantle cavity of submature specimens (DE 07) ciliated and provided with symmetrical arrangement of 11-15 lateral and dorsoterminal, stout and rather papilla-like respiratory organs ( Fig. 7A View FIG ). Th ese are about 60 µm long and have a 30-40 µm basal diameter, and are distally and laterally ciliated; each organ has two intruding muscle strands. Th is area is loosely overlain by suprapallial glands. In the two fully mature specimens (DE 05), however, only the terminal cavity shows – over a short distance – what appear to be free papillae ( Fig. 7B View FIG ). Th e epithelium of the latter, however, consists of large gland cells and no ciliation is visible; these gland cells are anteriorly continuous with the densely packed suprapallial glands (extending to the pericardium). These mature specimens, therefore, lack true respiratory organs. Central mantle cavity without lateral ciliation and forming a pair of lateral, dorso-ventrally oval pouchings ( Fig. 7 View FIG , pp); in the two fully mature specimens these are lined by 15-20 µm high epithelial glands (the “sac-formed glands” in Odhner 1921: 19, yet not really glandular in SMNH 4731). The ciliated rectum opens medio-dorsally. A strong, calcareous pallial hook with proximal excavation is differentiated at both sides of the anterior cavity opening ( Fig. 3 View FIG ).
The mantle cavity continues anteriorly to its opening, and the dorsal space forms a short, transversely separated (genital) pouch; the copulatory stylet sheaths open laterally. Ventrally, a characteristic, paired pouch or paired irregular sack opens; this paired sack extends anteriorly ( Figs 7 View FIG ; 9 View FIG , vp). The dorsal (genital) pouch is dorsally ciliated only; at its anterior end it receives, from ventral, the narrow opening of the central portion of the unpaired spawning duct ( Fig. 9 View FIG ).
Musculature
Body wall musculature inconspicuous, paired ventral reinforcement of longitudinal layer (used to roll up) distinct, wide. Dorso-ventral bundles moderate,
close to body wall. Strong and voluminous radula musculature; copulatory stylets with strong muscle bundles ( Fig. 8 View FIG ).
Sensory system
The unpaired cerebral ganglion (160-180 µm wide, 75 µm high) is slightly bilobed posteriorly and releases laterally the three pairs of connectives separately; two pairs of latero-frontal cerebral nerves begin with a distinct ganglionic swelling. Each (first) lateral ganglion is voluminous and located close to the cerebral ganglion. Elongate ventral ganglia (diam. 40 µm) behind the pedal pit interconnected by two commissures. Buccal ganglia (diam. 30-35 µm) lateral to opening of the pharynx into the midgut ( Fig. 4 View FIG ); their commissure runs between midgut and beginning radula sheath. The lateral cords are sparsely, the ventral cords more regularly provided with nuclei (medullar). Medullar suprarectal commissure rather short (60-80 µm × diam. 15-20 µm), positioned directly behind the anus.
Atrial (vestibular) sense organ short with densely arranged slender papillae (diam. 7-10 µm), basally united in groups of up to four. “Horseshoe-shaped” ciliary tract incomplete: it begins unpaired at midpoint of organ extension and then runs pairedly posterio-ventrally to end of the common atriobuccal opening; ventral tracts patchy, anteriorly missing.
Dorso-terminal sense organ prominent at dorsal end of body (behind end of mantle cavity; Fig. 7 View FIG ).
Alimentary tract
The foregut epithelium begins immediately adjacent to the atrial ciliary tract, dorso-posteriorly within the common atrio-buccal cavity. Due to longitudinal folding, the foregut epithelium (with moderate cuticle) appears to be papillate; it is loosely underlain by circular, longitudinal and radial muscles. Pharynx widened and strongly cuticularised, anterio-laterally pouched to house the distal portion of the strong lateral radula denticles. Anterior pharynx underlain by subepithelial, intercellularly opening glandular cells (pharyngeal glands), most densely laterodorsally; posterior pharynx with increased dorsal and lateral (radula-)musculature. Radula apparatus consists of sheath (400-450 µm) and paired, ventroposteriorly bending sack, which exceeds the sheath by about 150-300 µm ( Fig. 4 View FIG ; total length of radula apparatus in the investigated specimens therefore 500-750 µm); ventral sacks embedded in radula musculature along almost their entire length. In the sectioned specimens, the ventral sacks show an additional transverse leg (about 270°) distally in the U-shaped medial bend (of about 180°); the bends at each side are arranged above each other. Radula sheath paired for most of its extension towards pharynx, proximally unpaired (in cs U-shaped). Plates of biserial radula in pharynx 90-100 µm wide and 25-40 µm high, with a maximum of 15 denticles ( Fig. 5 View FIG ): a strong lateral denticle up to 170 µm long, 12 heterogeneous denticles at the distal border (not being precisely aligned and two of them possess a double cusp which is poorly visible in sections), and two small denticles somewhat below the border at the face ( Fig. 5 View FIG ). In distal ventral sacks, the radula plates measure 30-40 × 10 µm and still have about 15 rather homogeneous, small denticles. Radula support (bolster, odontophore) muscular, posteriorly with four pairs of differently sized turgescent cells.
Lateroventral foregut glandular organs are elongate-bulbous, surrounded by a thin lamina (reinforced basal membrane?) and embedded within radular musculature. Elongate gland cells epithelial, with main cell bodies longitudinally arranged (modified type C; cf. Todt 2006); secretion of large granula or droplets. Each organ distally with distinct central lumen; narrow duct (about 50 µm long) of pharyngeal epithelium opens laterally just anterior of pharynx transition into midgut ( Fig. 6 View FIG ).
Oesophagus lacking. Pharynx opens directly into midgut above beginning of the radula sheath.Midgut with short caecum, anteriorly paired. Dorsoventral muscle bundles run close to body wall, forming only weak midgut pouches. Middorsal ciliary tract continues in ciliation of hindgut. Posterior lumen of midgut and hindgut in one specimen of station DE 07 (paratype 1) was filled with an elongate, compact, non-cnidarian piece of prey.
Gonopericardial system
The two sectioned specimens of station DE 05 were fully mature. Gonad paired, not extending into anterior body; eggs up to 130 µm in diameter. Paired gonopericardioducts open frontally into spacious pericardium ( Fig. 7B View FIG ), with heart largely unattached. Pericardioducts emerge laterally; in fully mature animals, pericardium (filled with genital products) continues somewhat further posteriorly ( Fig. 7B View FIG ). Pericardioducts ciliated except medially, no vesiculae seminales; they run anteriorly, each opening into the spawning duct. Paired spawning ducts with tall, ciliated epithelium; in fully mature animals, behind opening of pericardioducts, each duct has a dorsal and ventral sacculation (probable receptacula seminis; Fig.7B View FIG ), posteriorly with increasing glandular contents. Junction of spawning ducts is not a simple fusion but a paired opening into an anteriorly blind-ending central portion; in cross section this yields for a short distance a characteristic, three-parted aspect ( Fig. 8 View FIG ). The united spawning ducts continue posteriorly as a voluminous, unpaired, glandular and ciliated organ, extending almost up to the (genital) pouch of the mantle cavity; here, only the central portion of this wide spawning duct opens from ventral into the pouch ( Fig. 9 View FIG ).
Paired copulatory stylet apparatus, each with two elements ( Figs 7-9 View FIG View FIG View FIG ) within a common sheath: 1) a very strong stylet that may extend (max. length 650 µm, max. diameter 50 µm) close to the beginning gonad. Th is stylet has a cavity along most of its length; the cavity is proximally wider and is filled by a tissue plug at the proximal end; and 2) a much shorter, entirely solid element (about 225 µm long) adjoining the stylet. It is proximally flattened and scale-like (up to 65 µm wide) and tapers distally to a point that ends earlier than the strong stylet. Sheath at each side surrounded (particularly at its proximal portion) by a strong envelope of musculature (protractors, retractors) up to 60 µm thick ( Fig. 8 View FIG ).
COMPARISONS
Based on the organization, the present specimens belong to the Simrothiellidae . Among the nine defined genera (see Scheltema & Schander 2000; Salvini-Plawen 2004), they must be classified within Simrothiella Pilsbry, 1898 . Herein, they are very similar to the type species Simrothiella margaritacea ( Koren & Danielssen, 1877) from Norway at 75- 115 m ( Odhner 1921; Salvini-Plawen 2004; not Scheltema & Schander 2000), particularly with respect to the hard parts (radula, etc.).
The differences between the two subadults and the two fully mature specimens pertain to respiratory organs and to the (probable) receptacula seminis. Other features of the secondary genital apparatus, however, are typically differentiated in the submature specimens, allowing determination to the species level (see also Salvini-Plawen & Paar-Gausch 2004). All four specimens of the new species differ in certain characters from S. margaritacea (see Odhner 1921; Salvini-Plawen 2004) and also from the specimens from off Galicia, Spain (cf. Todt 2006). These characters include: 1) the configuration of the mantle cavity with the differentiation of a paired ventro-anterior pouch or sack ( Figs 7 View FIG ; 9 View FIG , vp) and the peculiar condition of the respiratory organs ( Fig. 7 View FIG ; in S. margaritacea there is at most an unpaired ventro-anterior pouch and the gills show lateral foldings); 2) the denticulation at the distal border of the radula plates in adults (also contrasting the position of the denticles in S. abysseuropaea ; cf. Salvini-Plawen 2004), as well as the ventral radula sacks extending far behind (in S. margaritacea the sacks exceed the sheath by about 50-130 µm); 3) the presence of turgescent cells in the radula support or bolster (absent in S. margaritacea ); 4) the frontally paired midgut caecum (unpaired in S. margaritacea ); 5) the short supra-rectal commissure (in S. margaritacea up to 250 µm); and 6) the configuration of the spawn- ing ducts: before their junction they have a blind central portion into which the ducts open ( Fig. 8 View FIG ; in S. margaritacea the ducts simply fuse). These differences clearly define the new species.
These characters underline that solenogaster species should not be determined predominantly based on hard parts (Scheltema & Schander 2000). Specific characterisations using that approach fail in two respects: 1) hard parts are only superficially similar (see S. margaritacea vs. S. abysseuropaea in Scheltema & Schander 2000 and Salvini-Plawen 2004); and 2) hard parts (including the radula) are fairly identical in shape in species with different internal organization (present contribution).
KOREN J. & DANIELSSEN D. C. 1877. - Beskrivelse over nye arter, henhOrend til slaegten Solenopus, samt nogle oplysninger om dens organisation. Archiv for Mathematik og Naturvidenskab (Christiana) 2: 121 - 128 (in Norwegian, translated into English in 1879: Descriptions of new species belonging to the genus Solenopus, with some observations on their organisation. Annals and Magazine of Natural History ser. 5, 3 (17): 321 - 328).
ODHNER N. 1921. - Norwegian Solenogastres. Bergens Museums Aarbok 1918 - 1919 3: 1 - 86.
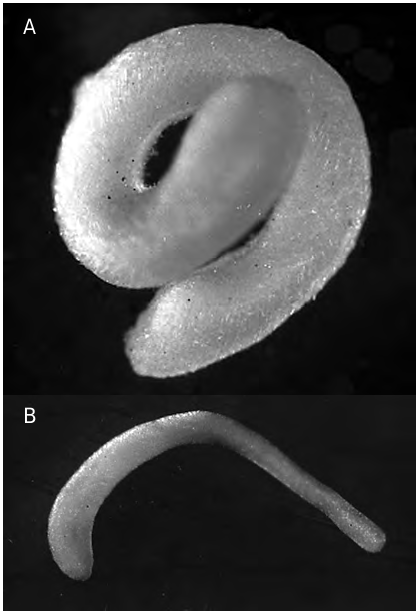
FIG. 1. — Simrothiella vasconiensis n. sp.:A, holotype in preserved state (posterior end below), 6.5 mm long; B, submature paratype 2 in preserved state (anterior end left), 5 mm long.
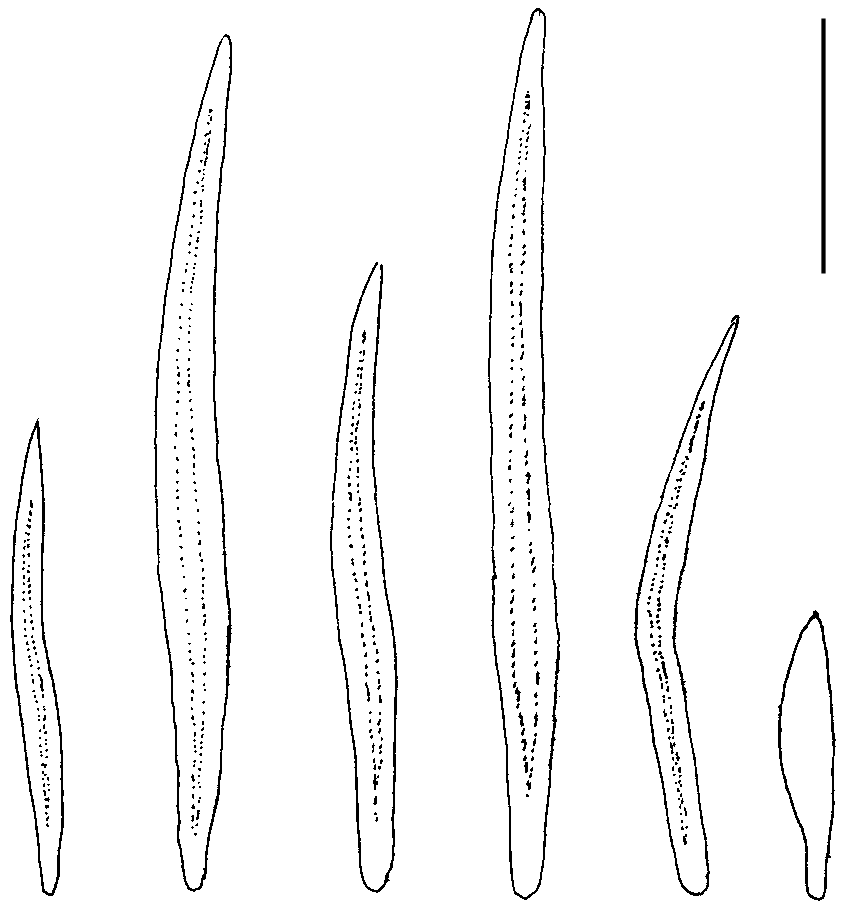
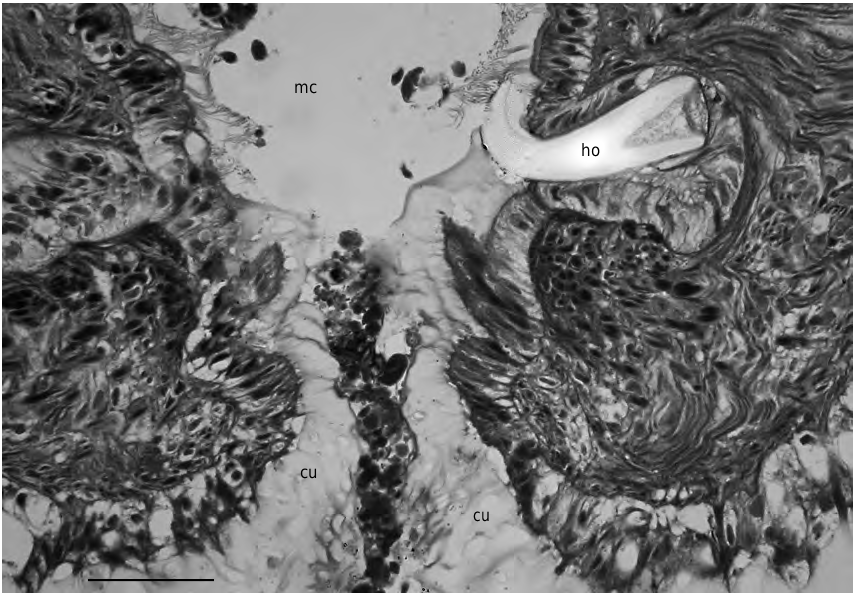
FIG. 3. — Simrothiella vasconiensis n. sp., cross section through beginning mantle cavity opening showing the calcareous pallial hook (paratype 2). Abbreviations: cu, mantle cuticle; ho, pallial hook; mc, mantle cavity. Scale bar: 25 μm.
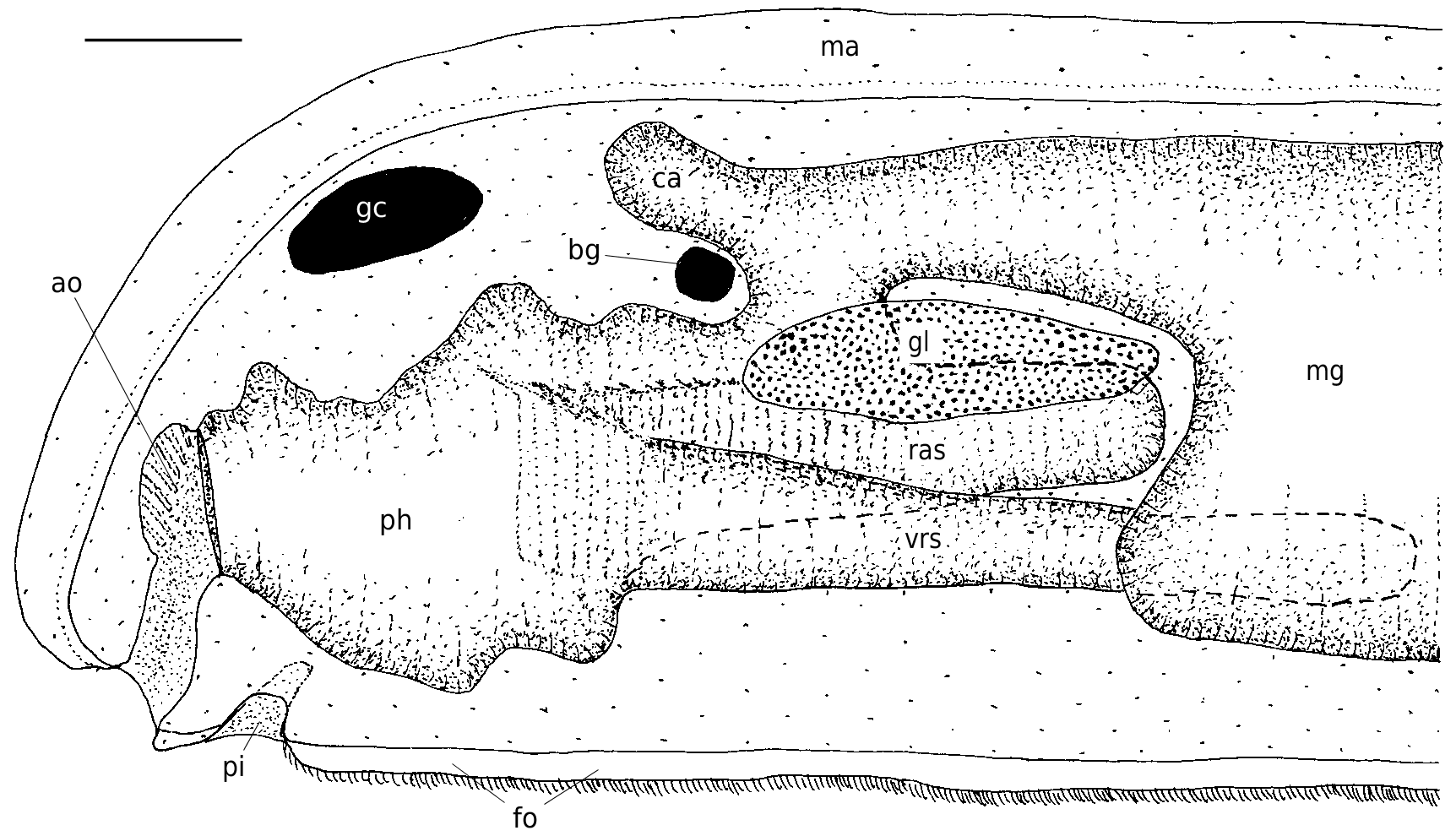
FIG. 4. — Simrothiella vasconiensis n. sp., organization of the anterior body viewed from left. Abbreviations: ao, atrial sense organ; bg, buccal ganglion;ca, midgut caecum;fo, pedal fold (foot);gc, cerebral ganglion;gl, lateroventral foregut glandular organ;ma, mantle; mg, midgut; ph, pharynx; pi, pedal pit; ras, radula sheath; vrs, ventral radula sack. Scale bar: 100 μm.
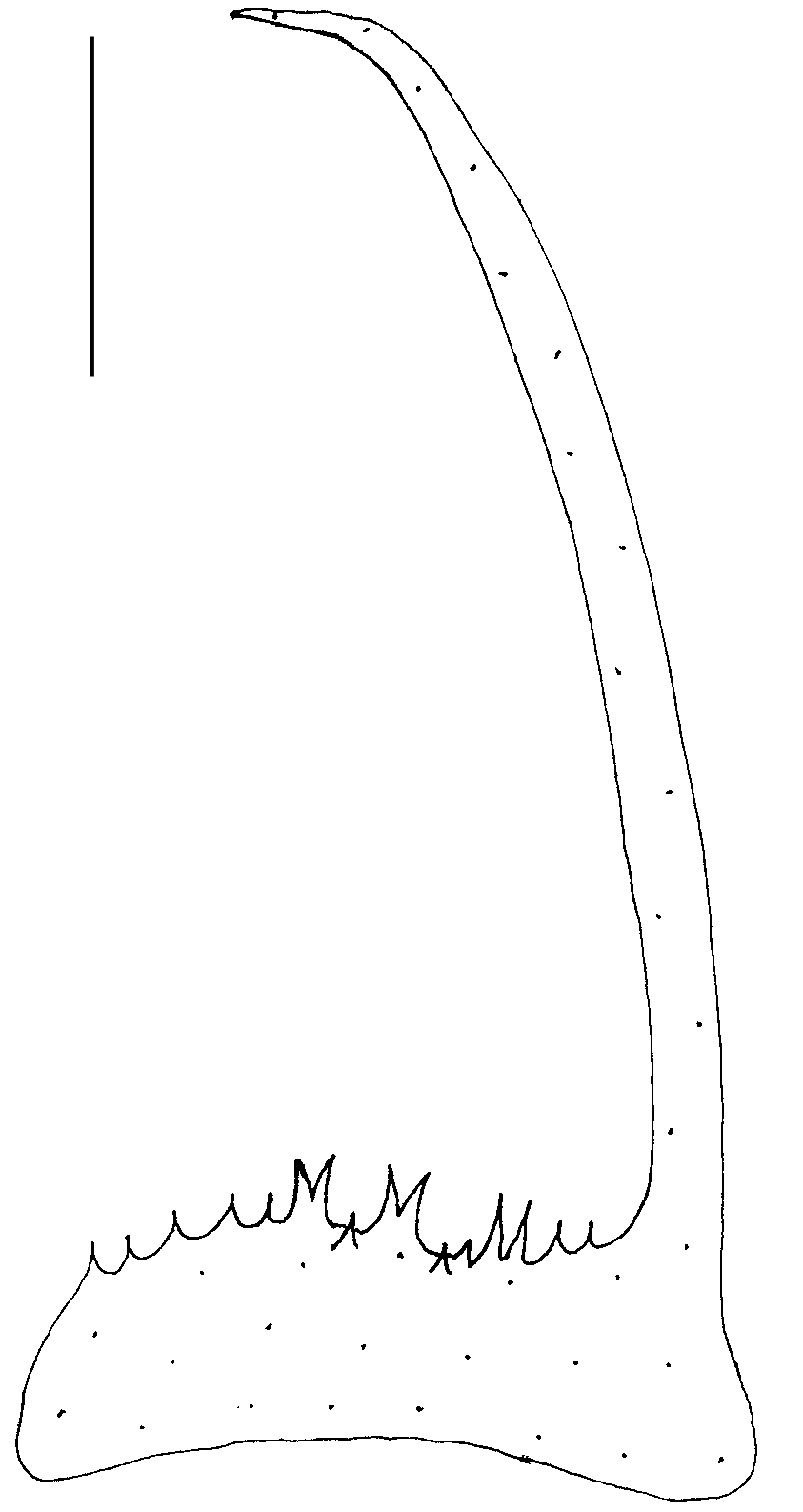
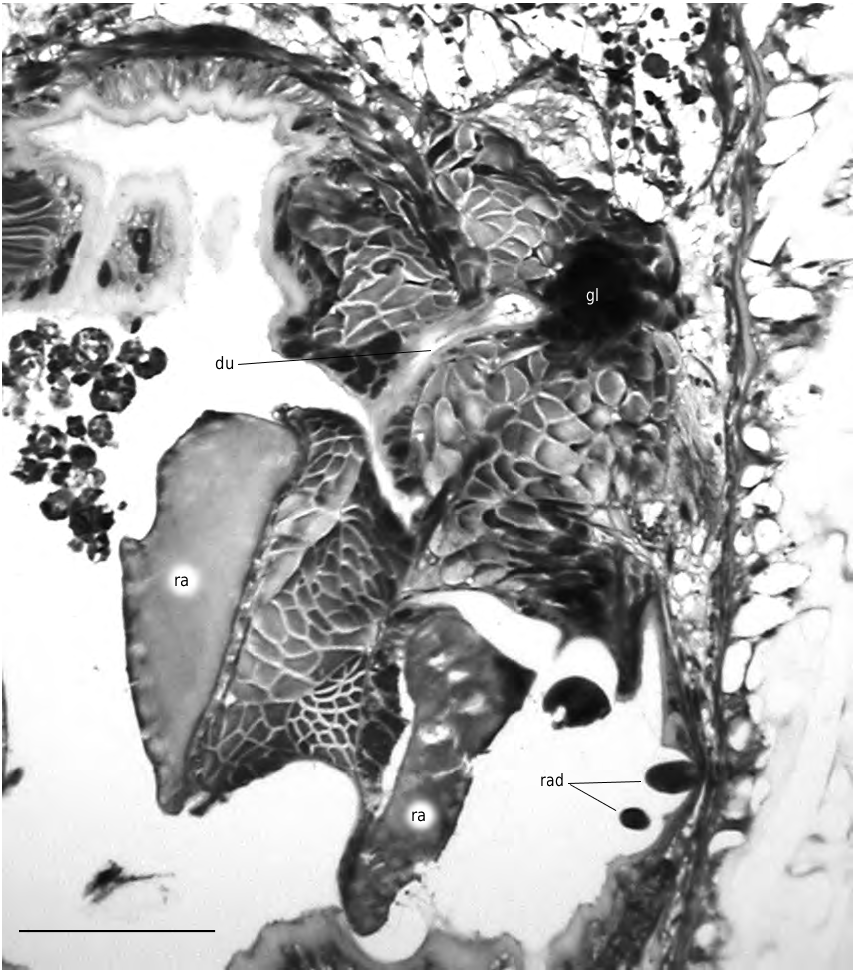
FIG. 6. — Simrothiella vasconiensis n. sp., right portion of cross section through pharynx just anterior of its transition into midgut with narrow duct of latero-ventral foregut glandular organ opening laterally (paratype 1). Abbreviations: du, outleading duct of foregut glands (gl); ra, radula plate; rad, cross sectioned long, lateral radula denticles. Scale bar: 50 μm.
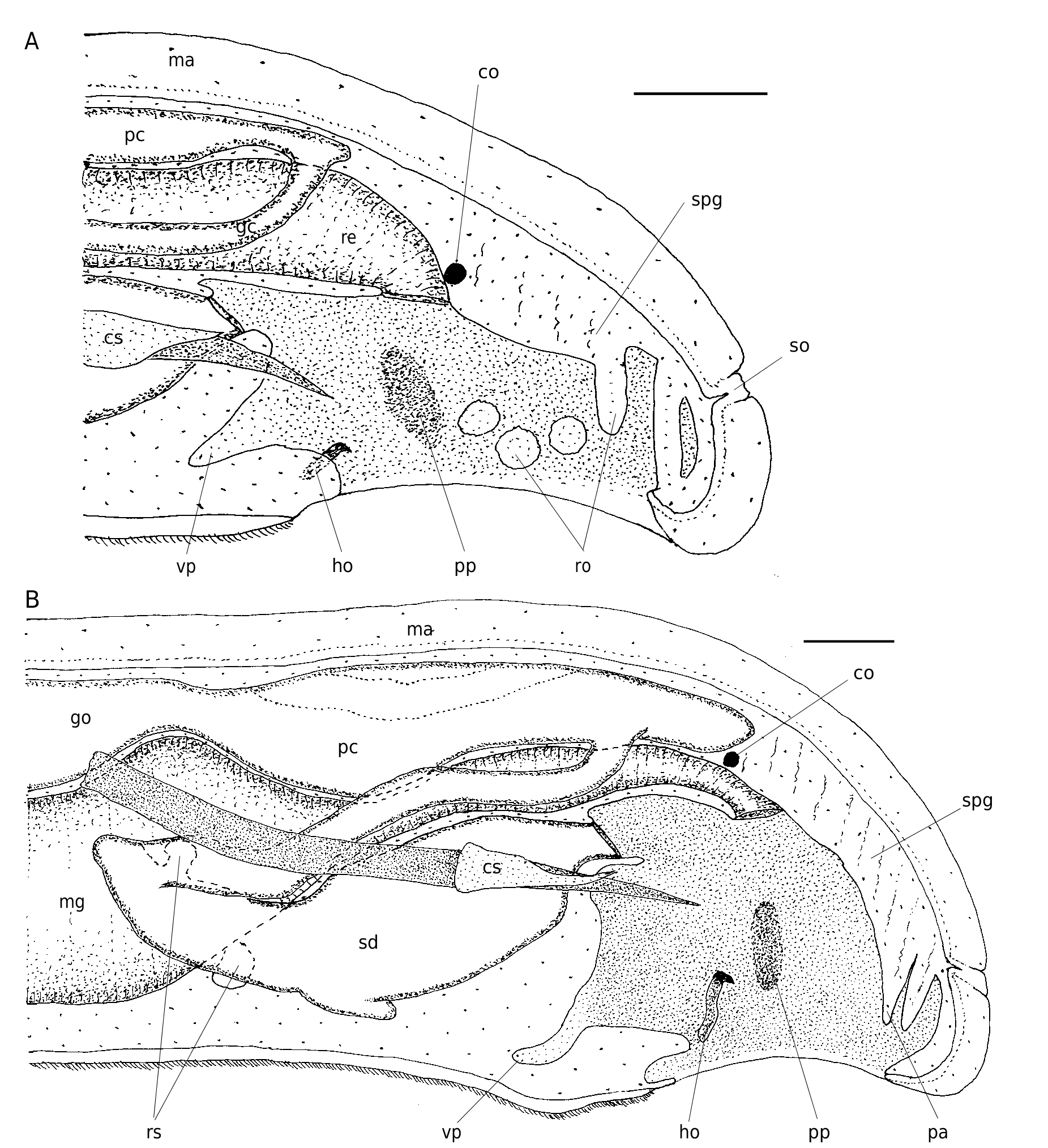
FIG. 7. — Simrothiella vasconiensis n. sp., organization of the posterior body: A, of a submature specimen viewed from left; B, of a fully mature specimen viewed from left.Abbreviations:co, suprarectal commissure;cs, copulatory stylet sheath;go, gonad;ho, pallial hook; ma, mantle; mg, midgut; pa, papillae; pc, pericardium (with heart); pp, lateral pallial pouch; re, rectum; ro, respiratory organs; rs, receptaculum seminis; sd, spawning duct; so, dorsoterminal sense organ; spg, suprapallial glands; vp, ventro-anterior pallial pouch. Scale bars: 100 μm.
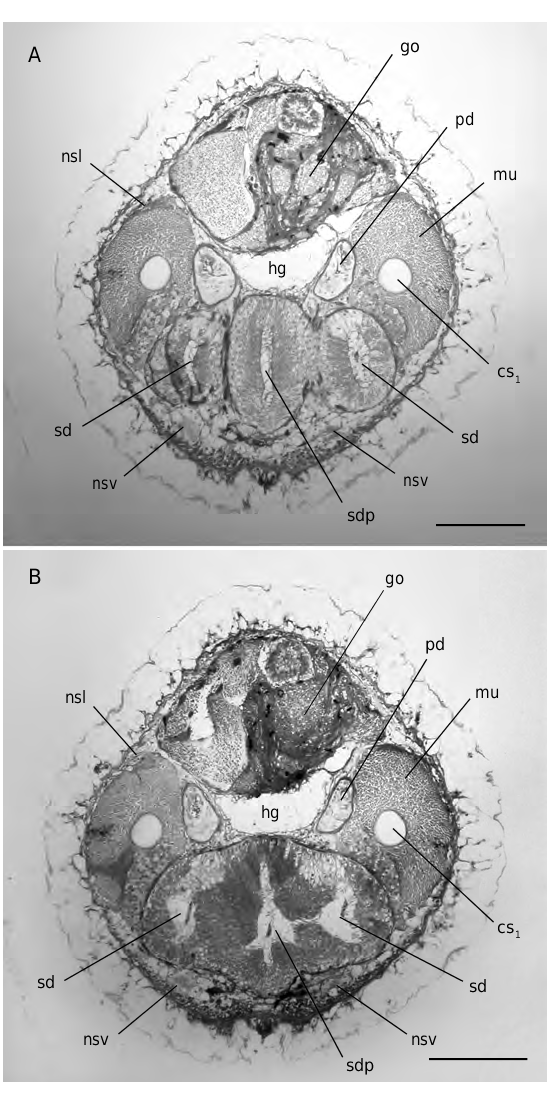
FIG. 8. — Simrothiella vasconiensis n.sp.,two cross sections through junction of spawning ducts (holotype):A, three-parted configuration with blind middle portion; B, fusion of spawning ducts with middle portion. Abbreviations: cs1, strong copulatory stylet; go, gonad; hg, hindgut; mu, musculature of copulatory stylets; nsl, lateral nerve cord; nsv, ventral nerve cord; pd, pericardio-duct; sd, spawning duct; sdp, blind anteriorly directed middle portion at junction of spawning ducts. Scale bars: 100 μm.
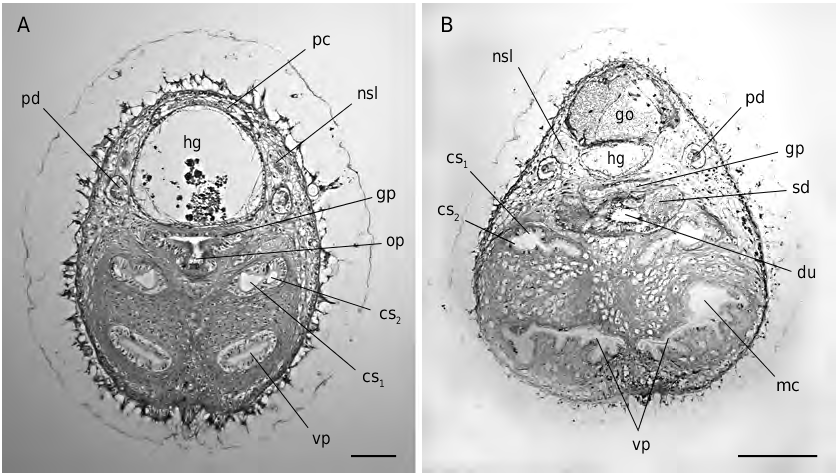
FIG. 9. — Simrothiella vasconiensis n. sp., cross sections through posterior body in the region of the outlet of the spawning duct: A, in the submature paratype 2; B, in the holotype. Abbreviations: cs1, strong, inner copulatory stylet; cs2, lateral, short copulatory stylet; du, central outlet (short duct) of spawning duct;go, gonad;gp, genital pouch of mantle cavity;hg, hindgut;mc, mantle cavity;nsl, lateral nerve cord; op, opening of central portion of spawning duct into genital pouch; pc, pericardium; pd, pericardioduct; sd, spawning duct; vp, paired ventro-anterior pouch of mantle (pallial) cavity. Scale bars: A, 50 μm; B, 100 μm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |