
Gen_Indet sp_indet, Molnar, 1978
|
publication ID |
https://doi.org/ 10.5281/zenodo.1039723 |
|
DOI |
https://doi.org/10.5281/zenodo.3483000 |
|
persistent identifier |
https://treatment.plazi.org/id/0392879F-D075-3E40-59A7-FBF7F8E8DAC8 |
|
treatment provided by |
Jeremy |
|
scientific name |
Gen_Indet sp_indet |
| status |
|
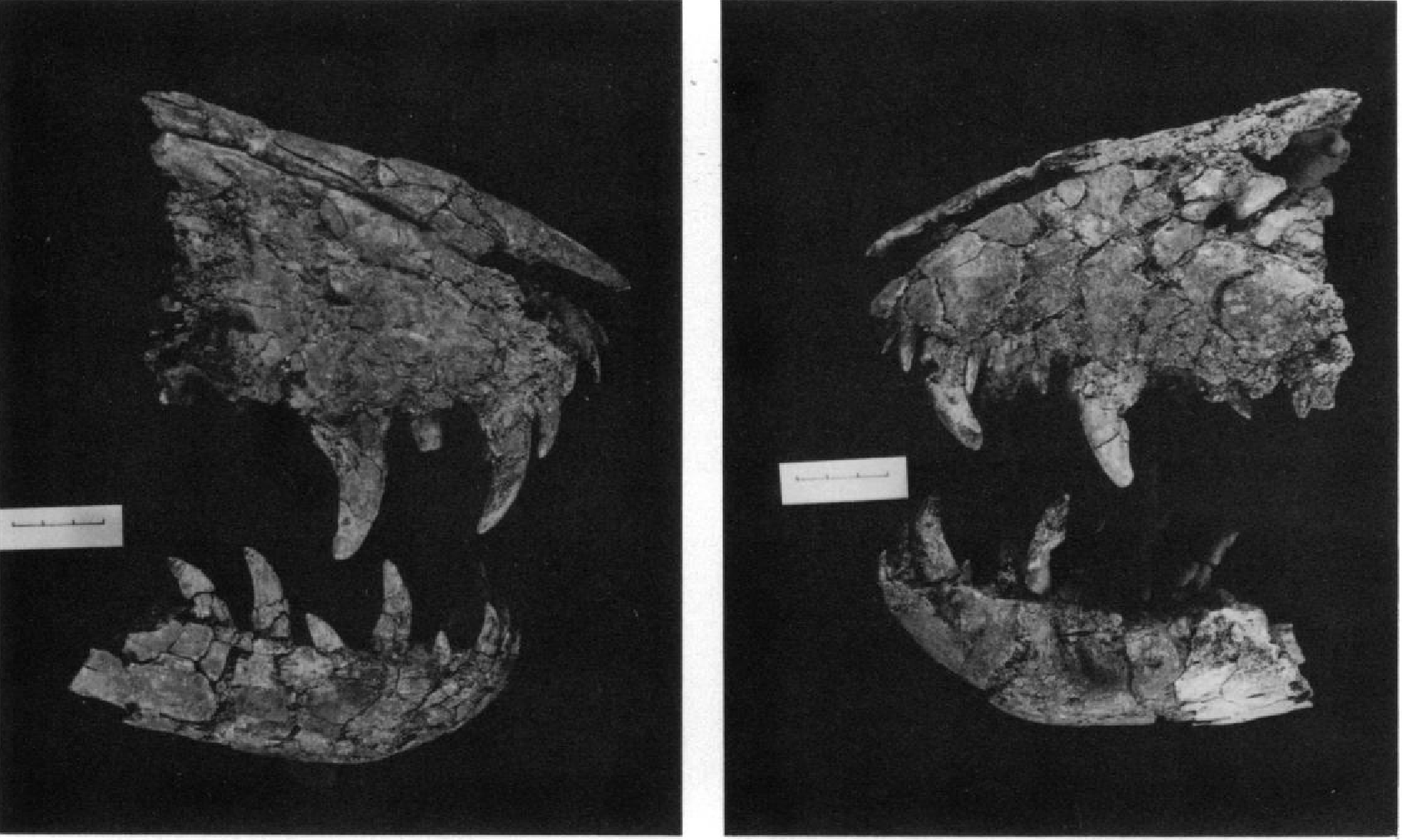
During the summer of 1966 Mr. Harley Garbani, of the Museum of Natural History, Los Angeles County, collected the snout and associated pieces of a theropod skull and jaws, catalogued as LACM 28471 View Materials . These were from a dark grey clay of the Hell Creek Formation on the F. S. McKeever Ranch, S34 T 21N R37E, near Jordan in Garfield County, Montana (LACM Locality 7245). The material was found near a large?Triceratops skull, but the level in the Hell Creek Formation was not recorded.
The specimen was initially identified as a juvenile Tyrannosaurus rex because of its small size and occurrence in the Hell Creek Formation. Thus the specimen is of considerable interest, for it represents an individual smaller than any known to pertain to the Tyrannosauridae. However, it appears likely to be neither Tyrannosaurus rex nor Albertosaurus lancensis (although possibly juvenile) and the existence of a third large theropod in the Hell Creek is itself of interest. Finally, it is one of the few theropods from which an endocranial mold has been prepared. Although the specimen is not referrable to any well known taxon, the incompleteness of the material renders it unwise to erect a new taxon for it at this time,
and following Galton (1973) in a similar situation, it will herein be called the Jordan theropod .
COLLECTION DESIGNATIONS
AMNH — American Museum of Natural History (New York).
LACM — Museum of Natural History, Los Angeles County (Los Angeles).
MMS — Minnesota Museum of Science (St. Paul).
NMC — National Museum of Natural Sci ences (Ottawa).
PIN — Paleontological Institute of the Academy of Sciences of the USSR. (Moscow).
USNM — United States National Museum (Washington).
YPM — Peabody Museum of Natural History, Yale University (New Haven).
DESCRIPTION
Maxilla.— Both maxillae ( Text-fig. 1 View Text-fig. 1 ) are preserved posteriorly to a line just behind the anterior margin of the antorbital recess. The term antorbital recess is used here and throughout to designate the excavation often surrounding the antorbital fenestra(e) in archosaurs. Similarly the term supratemporal recess will be used for the excavation sometimes surrounding the supratemporal fenestra. The anterodorsal and ventral margins of the maxilla converge anteriorly at an angle of 50 degrees, as compared to 80 in Tyrannosaurus and 70 (in immature specimens) to 90 in Albertosaurus . This, together with the shallow nasals, gives the preserved portion of the snout a more nearly triangular appearance than is usually found among Cretaceous theropods. Anteriorly there is a row of foramina 1 cm above the ventral margin.
Little of the antorbital recess is preserved. Its anterodorsal margin slightly overhangs the lateral wall and a foramen extends anterodorsally from the anterior apex of the recess. This would correspond with the anteriormost antorbital fenestra of Russell (1970). It is clear that were a second (anterior) antorbital fenestra present, it did not extend to the anterior margin of the recess.
A shallow concavity in the anterodorsal margin of the left maxilla marks either the ventral margin of the naris or of the subnarial foramen (sensu Madsen, 1976A). This concavity is 3.5 cm long and terminates anteriorly 0.5 cm above the ventral margin.
The medial surfaces of both maxillae have been obliterated, except for two medially directed shelves subparallel to the ventral margin. The anterior shelf is just ventral to the posterior termination of the previously described concavity 3 cm above the alveolar margin. This process is unbroken at its posterior termination as is the posterior shelf at its anterior termination. This suggests the existence of a channel between the two, possibly similar to the internal passage connecting to the subnarial foramen in Allosaurus ( Madsen, 1976A). However, in Allosaurus this passage opens between the maxilla and premaxilla not within the maxilla. Both shelves (each 2 cm long as preserved) slope downwards anteriorly. They are presumably portions of the palatal process of the maxilla.
The form of the maxilla reconstructed to correct for displacements at the breaks is given in Text-figure 2 View Text-fig. 2 .
Nasal.— Both nasals ( Text-fig. 1 View Text-fig. 1 ) are preserved for a length of 16 cm posteriorly from the premaxillary junction. The maximum width, near the posterior end of the nares, is at least 3.4 cm. The nasals preserved in articulation are long, narrow, parallel-sided elements slightly flared at the maxillary processes. Strongly arched in the transverse plane, their maximum height is 1.4 cm. The premaxillary processes diverge anteriorly to receive between them the nasal processes of the premaxillae for a distance of 1 cm.
The dorsal surface of the nasals is smooth: the median contact is preserved anteriorly but apparently obliterated by fusion beyond a break 5 cm from the premaxillary contact. The naso-maxillary suture is smooth as in dromaeosaurids and unlike the dentate appearance it assumes in tyrannosaurids.
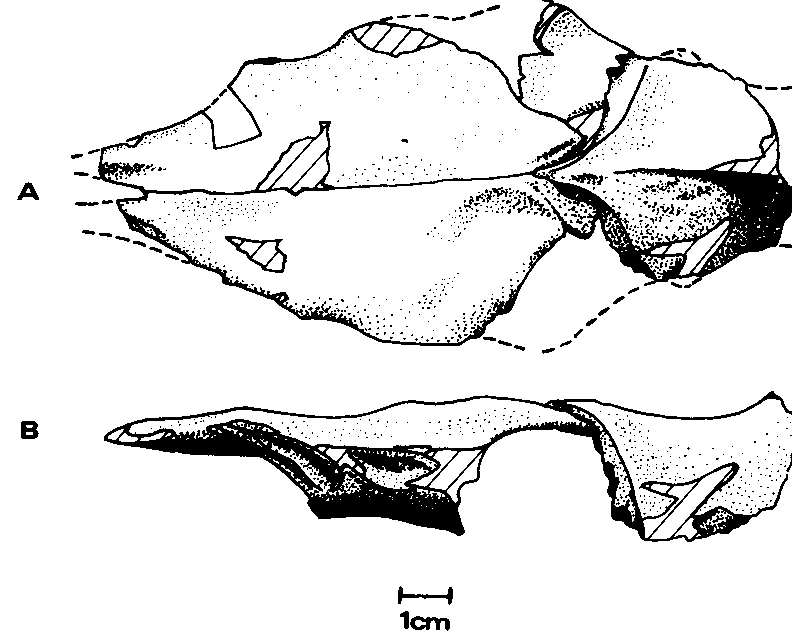
Frontal.— The right frontal ( Text-fig. 3 View Text-fig. 3 ) lacks only the lateral portion (contacting the postorbital) while the left is less complete. The frontal is subtriangular in form when viewed dorsally, with the most acute angle directed anteriorly, an oblique angle directed posteromedially, and the third, truncated, directed posterolaterally.
The fronto-parietal contact is V-shaped, with the apex directed anteriorly, and the sutures curving laterally until they are in the transverse plane. This contact is a serrate suture, and thus immobile, in contrast to that of Dromaeosaurus albertensis ( Colbert and Russell, 1969) . The posterolateral region of the right frontal shows a serrate sutural laterosphenoid contact surface. The postorbital contact, preserved on the left frontal, is a shallow, anteroposteriorly elongate concavity approaching to within 3 mm of the prefrontal contact. Since the surface here is worn it is not clear whether the postorbital came into contact with the prefrontal thus excluding the frontal from the orbital margin. Both prefrontal contact surfaces are preserved. They are deep, teardrop-shaped concavities, with the “tails” of the “teardrops” directed anterodorsally. Ventrally adjacent to the “tail” is a deep groove. The articular surface for the nasal, although worn, shows a medial trough presum- ably occupied by a tongue of the nasal. Lateral shelves were presumably also occupied by portions of the nasals which thus slightly overlapped the frontals at the junction, and gave the contact the shape of a W when viewed from above. Such a contact is also found among the tyrannosaurids (e.g. Albertosaurus , Daspletosaurus', Russell, 1970).
The dorsal surface of the frontals is smooth rising posteriorly to the parietal junction. This rise extends 1.5 cm anteriorly from the contact along the midline to form a low sagittal crest. There is no indication of invasion of the frontals by a supratemporal recess, and although this portion of the surface is worn, the wear is so slight as to obscure none but the faintest indication of such a recess. Hence the frontals differ from those of many other theropods { Allosaurus , Dromaeosaurus , the saurornithoidids, the tyrannosaurids, etc.) in which the recesses unmistakably invade them.
The ventral surface of the frontals is better preserved and from this a partial endocranial mold has been made. The cristae cranii, the longitudinal ridges bounding the endocranial cavity, are the most prominent features. Only the anterior portions of the cristae are preserved. They show no sign of union with a bony element ventrally in the region of the taenia medialis, unlike the case in Allosaurus ( Madsen, 1976A), Saurornithoides ( Barsbold, 1974) , Tyrannosaurus ( Osborn, 1912; Bellairs, 1949), and a specimen tentatively referred to Albertosaurus (LACM 23845). In the Jordan theropod the cristae seemingly contacted only epioptic membrane. The cristae are slightly curved, convex medially, giving a constriction of the olfactory passage as in the tyrannosaurids and Dromaeosaurus .
Parietal.— Only the anteromedial portion (5 cm in length) of the parietals ( Text-fig. 3 View Text-fig. 3 ) is preserved, fused along the midline. The maximum width as preserved is 4.2 cm, which since portions of both proötic contacts are present, must be close to the actual value. The proötic union was a serrate suture.
The smooth dorsal surface rises medially to a sagittal ridge which, in turn, rises abruptly posteriorly suggesting a supraoccipital crest was present (but not preserved). The line of intersection between the ventral surface and the sagittal plane is smoothly concave ventrally, with only a small depression on the midline near the center of the portion preserved.
Dentary.— Anterior portions of both dentaries (Text-fig. 1) are preserved with the symphysis in contact. The symphysis is 4 cm long, the preserved portion of the right dentary 15 cm long, and that of the left 14.5 cm long. Both are 4 cm deep and 1.8 cm wide at the posterior break. While resembling those of most theropods, these dentaries differ in two characters: 1) the ventral margin of the symphysis is less inclined in lateral aspect, 40 degrees to the horizontal, versus 50 degrees in mature tyrannosaurids ( Albertosaurus , Daspletosaurus and Tyrannosaurus ) and 2) the alveolar margin ascends anteriorly, then levels off at the region of the symphysis like that of Baurusuchus ( Romer, 1956, Fig. 75D). Only Kelmayisaurus petrolicus ( Dong, 1973) and the immature Albertosaurus libratus (AMNH 5664) show a comparably low angle to the symphysis. Spinosaurus ( Stromer, 1915) and Marshosaurus ( Madsen, 1976B) alone among the theropods show such a rise, although some (e.g. Tyrannosaurus ) exhibit a downcurving of the ventral margin toward the symphysis.
The smooth medial and lateral surfaces of the dentaries are reasonably well preserved and show neither median longitudinal bars nor interdental plates. On the lateral surface a row of foramina lies 1.5 cm below the dorsal margin posteriorly.
Surangular.— A fragment 5 cm long of this element is preserved ( Text-fig. 4 View Text-fig. 4 ). Comprised are a portion of the articular facet for the quadrate, the facet for insertion of the M. adductor mandibulae externis superficialis, and a small portion of the lateral surface. Because unbroken surface can be traced in a transverse plane all around the piece just anterior to the quadrate contact it is probable that a surangular foramen was present. A lateral buttress lay just above the foramen suggesting the presence of a lateral fossa as in the dromaeosaurids ( Ostrom, 1969).
Teeth.— Six nearly complete crowns are preserved in the two maxillae ( Text-fig. 5 View Text-fig. 5 ). These are the first (1.6 cm in length), second (4.3 cm), and fourth (4.7 cm) right, and the first (1.8 cm), second (3.5 cm), and fourth (4 cm) left. The third on each side is represented by a partially erupted crown. All maxillary teeth are similar in form and show 15 serrations per 5 mm along the anterior carina, and 12 to 18 per 5 mm along the posterior. Four of these teeth lay anterior to the antorbital recess. The first shows no wear, and the crowns of the other maxillary teeth are uniformly 2.5 times as long as that of the first.
A tooth, adhering to the left maxilla, of different form from the maxillary teeth is assumed to be a premaxillary tooth. The crown is 1.2 cm long, shorter than those of the maxillary teeth, and is similar in form to the premaxillary teeth of tyrannosaurids and Dromaeosaurus with carinae at the posterolateral and posteromedial corners. The tooth is D-shaped in cross section and differs from Tyrannosaurus premaxillary teeth in being shorter anteroposteriorly and in lacking serrations.
All but the first dentary teeth are similar to those of the maxilla, but smaller. The largest preserved is 3.2 cm high, and although another appears larger (3.7 cm) this is due to slippage partially out of the alveolus. The first dentary tooth (crown 0.9 cm long) is smaller than the second, and its anterior carina is displaced medioposteriorly, the serrations running along the medial surface of the tooth. None of the dentary teeth are D-shaped in cross section. The first tooth resembles the symphyseal premaxillary teeth of Deinonychus antirrhopus ( Ostrom, 1969, fig. 24D) and the first dentary tooth of Dromaeosaurus . The second dentary tooth (1.9 cm high) is smaller than the succeeding teeth which range from 2.4 to 3.2 cm in length of crown. The more posterior of the dentary teeth show 15 to 20 serrations per 5 mm along the anterior carina, and 14 to 15 along the posterior: those of the first tooth are too worn for a count.
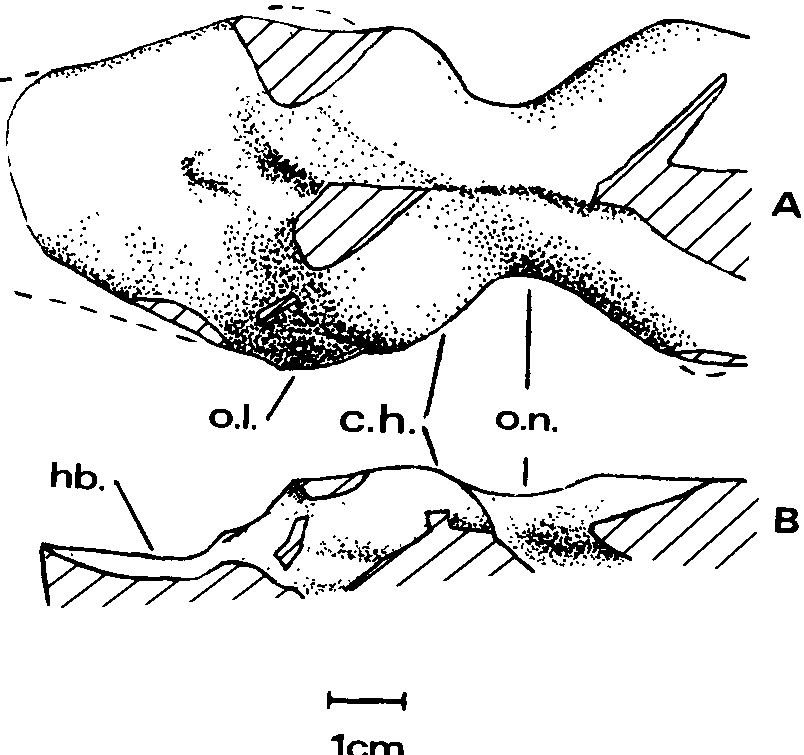
Endocranial mold.— The partial mold ( Textfig. 6 View Text-fig. 6 ) has been prepared from the frontals and parietals. Just anterior to the cerebral hemispheres is a constriction of the olfactory passage, which then abruptly widens anteriorly. Such a constriction is also present in the olfactory passages of Allosaurus and Tyrannosaurus ( Osborn, 1912) , Tarbosaurus ( Maleyev, 1965) and Ceratosaurus ( Marsh, 1896) . In both Tarbosaurus and Tyrannosaurus the constriction is less abrupt than in the Jordan theropod, which more resembles Ceratosaurus in this respect.
The cerebral hemispheres are prominent, bulging both dorsally and laterally, but not as much enlarged as in Stenonychosaurus ( Russell, 1969, fig. 3C). A slight expansion of the right side posteroventral to the cerebral hemisphere probably represents the right optic lobe, which was relatively larger than in Allosaurus , Stenonychosaurus or Tyrannosaurus , more like that of Tarbosaurus ( Maleyev, 1965) . The mid- and hindbrain region is distinctly lower than the forebrain region and flatter. This is in marked contrast to the conditions in Tarbosaurus and Tyrannosaurus where there is a slope upward to peak beneath the supraoccipital crest and then downward toward the foramen magnum. In the Jordan theropod the crest was considerably to the posterior of this peak of the endocranial cavity, giving the profile an appearance different from those of the tyrannosaurids. The descending posterior surface of the forebrain region exhibits a small projection (corresponding to the previously described pit of the parietals) similar to, but smaller than, that shown by the endocranial mold of Dromiceiomimus brevitertius ( Russell, 1972) . It presumably housed a parietal organ.
Nerve foramina are not evident on the mold (the region preserved being dorsal to them), nor are there any dorsal channels opening from the endocranial cavity as in Tyrannosaurus (but not Tarbosaurus ).
DISCUSSION
Maturity of the specimen.— The size of the Jordan theropod suggests that it might represent an immature tyrannosaurid. Recent work ( Rozhdestvenskii, 1965; Dodson, 1975B) indicates that immature dinosaurs are often referred to new taxa rather than recognized as immature. The best method of showing the immaturity of a given specimen is the presentation of a growth series, as is known for Allosaurus ( Madsen, 1976A) and Tarbosaurus ( Rozhdestvenskii, 1965) . Lacking such a series, one may extrapolate from known ontogenetic trends of the skeleton or look for features taken to be indicative of immaturity. Neither of these latter methods is conclusive for there is no certain way of identifying an immature individual as such from a single skeleton, given the continuous growth (implying open sutures) and the usual lack of terminal epiphyseal bones of reptiles ( Dodson, 1975A).
In the absence of a growth series characters of the Jordan theropod must be compared with predictions from known ontogenetic trends. As few such trends have been inferred in theropods, those of crocodilians (chosen because they exhibit closer phylogenetic relationship to theropods than other living reptiles and a more conservative structure than birds) are used.
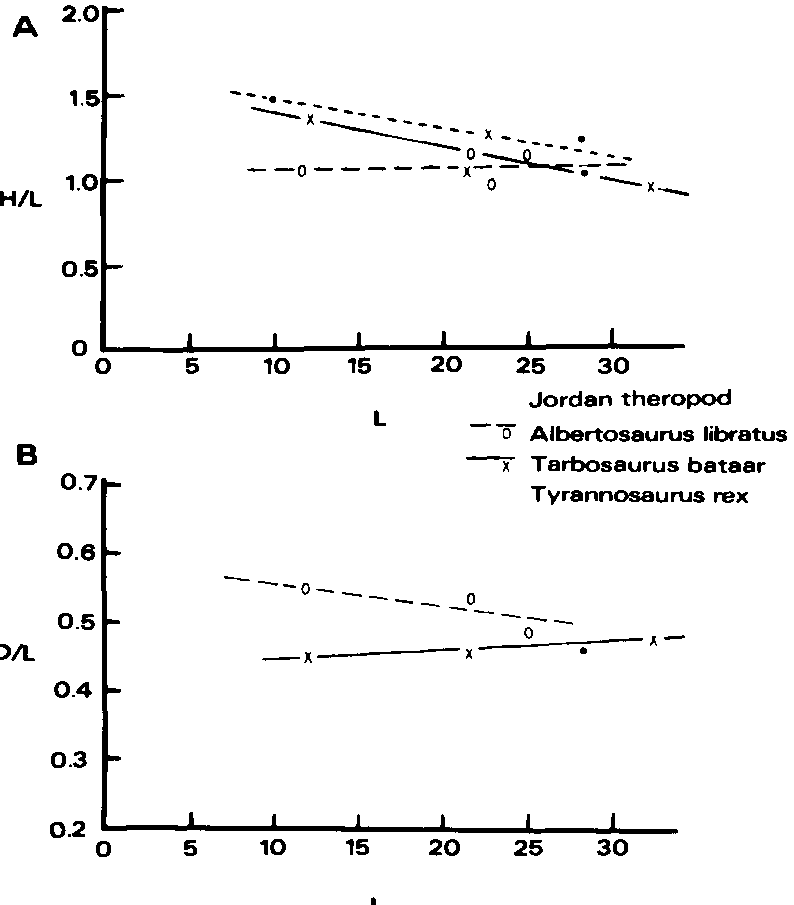
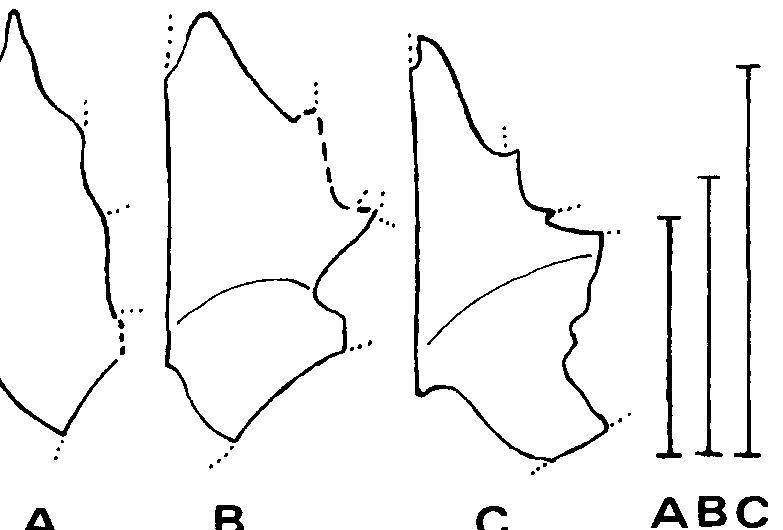
Crocodilians exhibit three trends of interest here: increase in relative length of the snout with age, decrease in separation between the supratemporal fenestrae with age, and increase in depth of dentaries with age ( Dodson, 1975A; Kalin, 1933, 1955). Similar trends have been indicated in lambeosaurs ( Dodson, 1975B). The first trend can also be demonstrated for tyrannosaurids, in Tarbosaurus and Tyrannosaurus ( Text-fig. 7 View Text-fig. 7 ). In Albertosaurus libratus the smallest skull, of AMNH 5664, has suffered dorsoventral crushing of the snout (Gilmore, 1945) which may conceal such a trend. The preserved portion of the snout of the Jordan theropod is longer relative to its height than that of any tyrannosaurid ( Textfig. 7 View Text-fig. 7 ), suggesting that it is not an immature tyrannosaurid.
The parietals of the Jordan theropod indicate the supratemporal fenestrae were more widely separated than in mature tyrannosaurids, suggesting that it may have been immature (or small).
The dentary of the Jordan theropod is shallower with respect to the length of the maxilla (anterior to the antorbital recess) than in any tyrannosaurid ( Text-fig. 7 View Text-fig. 7 ), but is most closely approached by that of PIN 552-2, an immature Tarbosaurus bataar (the type of “ Gorgosaurus novojilovi ”). This also suggests that the Jordan theropod was immature (or small).
Other features consonant with the interpretation of immaturity are: the smooth nasals, the lack of ossification ventral to the cristae cranii, the elongate form of the frontals ( Textfig. 8 View Text-fig. 8 ) and possibly the smooth naso-maxillary union. Yet all of these can also be found in adults of smaller taxa.
Several authors (e.g. Ostrom, 1969; Colbert and Russell, 1969) have used serration count as a diagnostic character of theropods. The number of serrations per 5 mm of crown is greater in the smaller genera ( Ostrom, 1969, Table 2). As far as I know only Gallup (pers. comm., 1975) has tested this trend, with isolated teeth of different sizes attributed to Acrocanthosaurus . He has found that smaller teeth have the greater serration count. Although this suggests that the serration count may change in ontogeny, other explanations are possible and more work on this point is desirable. The Jordan theropod shows a higher serration count than Albertosaurus and Tyrannosaurus ( Ostrom, 1969) , possibly consonant with interpretation of immaturity, if Gallup’s trend is substantiated.
These considerations suggest, but do not prove, that the Jordan theropod may have been immature.
Taxonomic affinities.— The Jordan theropod shares several characters with mature tyrannosaurids: the surangular foramen and buttress, D-shaped premaxillary teeth, and W-shaped naso-frontal contact. Interpretation of it as an immature tyrannosaurid allows accommodation of a suite of characters in which it differs from adult tyrannosaurids such as the smooth nasals, acute anterior angle of the maxilla, shallow dentary and shallow angle of the symphysis (all found in AMNH 5664). But other characters cannot be accommodated.
Comparison with the two contemporaneous well known tyrannosaurids, Albertosaurus lancensis and Tyrannosaurus rex , shows that both have a deeper maxilla than the Jordan theropod ( Text-fig. 9 View Text-fig. 9 ). This is especially true of the juvenile maxilla of T. rex recently described by Lawson (1975), which also differs from the Jordan theropod in that the maxillary fenestra is adjacent to the anterior margin of the antorbital recess in T. rex . The most similar tyrannosaurid maxilla is that of AMNH 5664, the immature Albertosaurus libratus . Even though this maxilla has suffered dorsoventral crushing (of which there is no indication in the Jordan theropod) it is still higher than that of the Jordan theropod.
Compared to those of the Jordan theropod, the frontals of tyrannosaurids are relatively thick (cf. Osborn, 1912, Pl. III). Admittedly immature tyrannosaurids undoubtedly bad thinner frontals than adults, but those of LACM 23845 (tentatively attributed to A. lancensis ) are 1.4 times the length of those of the Jordan theropod, but 2.5 times the thickness.
Lack of invasion of the frontals by the supratemporal recesses may indicate immaturity, but invasion is present in AMNH 5664, and lacking in several large theropods, e.g. Chilantaisaurus ( Hu, 1964) , Indosaurus (v. Huene and Matley, 1933) and Labocania ( Molnar, 1974) .
The postorbital contact of the frontal is basically similar in the four well known tyrannosaurid genera, subcircular in form and interlocking. This is quite different from the simple, anteroposteriorly elongate concavity of the Jordan theropod.
The form of the prefrontal contact of the frontal in tyrannosaurids is not described in the literature, but two specimens, LACM 23845 and MMS 51-2004 tyrannosaurus sp.), exhibit this surface. In both it is dorsoventrally, not anteroposteriorly, elongate and in MMS 51-2004, an interlocking suture not a simple concavity.
Taken together these characters indicate that the Jordan theropod, while showing a superficial resemblance to an immature tyrannosaurid, cannot reasonably be assigned to any known tyrannosaurid genus. Of the other late Cretaceous theropods, many, e.g. Alectrosaurus, Chingkankousaurus, Coelosaurus, Dryptosaurus, Macrophalangia, Pneumatoarthrus lack material in common with the Jordan theropod and cannot be compared. Others, the ornithomimids, saurornithoidids and oviraptorids, are obviously different. The dromaeosaurids are the most similar.
Resemblances of the Jordan theropod to dromaeosaurids include the surangular foramen and buttress, smooth and thin parallelsided nasals, absence of interdental plates in the dentaries (present in tyrannosaurids), serration count, shallow mandibles, thin frontals and lack of D-shaped anterior dentary teeth.
The forms of the prefrontal and postorbital contacts of the frontals of dromaeosaurids are poorly known. Figures in Colbert and Russell (1969) indicate that the postorbital-frontal junction of Dromaeosaurus albertensis , like that of the Jordan theropod, was anteroposteriorly elongate.
Colbert and Russell also comment that the braincase of Dromaeosaurus was more broadly open anteroventrally than in many theropods, and this too is indicated for the Jordan theropod.
D-shaped premaxillary teeth, however, are not common in dromaeosaurids ( Ostrom, 1969), Dromaeosaurus itself being an exception. Thus this should offer no bar to consideration of the Jordan theropod a dromaeosaurid. It should be noted that such premaxillary teeth are widespread among theropods, in addition to occurring in tyrannosaurids and Dromaeosaurus , they also occur in Allosaurus ( Madsen, 1976A), Ornitholestes ( Matthew and Brown, 1922) and have been attributed to Elaphrosaurus ( Stromer, 1934) . Such teeth are obviously adaptive in grasping and holding prey and may have been derived independently in different groups. Their use as an important taxonomic character stands in doubt.
Thus the Jordan theropod shares with dromaeosaurids five characters not shared with tyrannosaurids, but with tyrannosaurids only one character (the W-shaped naso-frontal contact) not shared with dromaeosaurids. The Jordan theropod may be best considered an exceptionally large dromaeosaur.
Implications.— The Jordan theropod material is too fragmentary for any reconstruction or inference of life habits beyond the obvious. The endocranial mold suggests well developed optic lobes, and hence a more prominent role of vision than in many theropods. The olfactory passage is wider relative to forebrain width than in Ceratosaurus or Stenonychosaurus , more like the condition in Allosaurus and the tyrannosaurids. This suggests a prominent role for olfaction (although not especially prominent relative to reptiles in general, cf. Jerison, 1973). The skull of the Jordan theropod lacks a mesokinetic hinge and shows no indication of other types of kinesis, and, if as Gow (1972) has suggested, cranial kinesis is an adaptation for feeding on prey relatively large with respect to the predator, this suggests that the Jordan theropod fed upon prey small with respect to its own size. The supratemporal recess is probably associated with development of the jaw adductors, especially the M. adductor mandibulae externis group and M. pseudotemporalis. Absence of the recesses in the Jordan theropod suggests weaker development of these muscles than in, e.g., the tyrannosaurids, and again a relatively small prey size.
Rosenzweig (1966) introduced for mammalian carnivores the concept of hunting set: a group of similar (often related) sympatric species which share similar hunting strategies. Such a set is permitted by the selection of different prey sizes by the different members, often reflected in their size. It is not the intent here to discuss the existence of such sets among theropods, but to point out that the skulls of Hell Creek theropods can be arranged in a sequence of increasing size approximating a hunting set ( Text-fig. 10 View Text-fig. 10 ). Interesting also is that unless the skull of the Jordan theropod was remarkably long and low it fills a gap in this sequence.
Finally the Jordan theropod and other recently described theropods ( Welles, 1970, 1972; Madsen, 1974, 1976B; Molnar, 1974) clearly point up the incompleteness of our knowledge of Mesozoic tetrapod faunae, even where large forms and ostensibly well known communities, such as those of the Morrison and Hell Creek Formations, are concerned.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.