
Calycopis janeirica
|
publication ID |
https://doi.org/ 10.5281/zenodo.192050 |
|
DOI |
https://doi.org/10.5281/zenodo.5627400 |
|
persistent identifier |
https://treatment.plazi.org/id/038EE54B-FFB9-FF81-FF5C-FBDDFBA081BA |
|
treatment provided by |
Plazi |
|
scientific name |
Calycopis janeirica |
| status |
|
Calycopis janeirica View in CoL (Felder, [1863]) ( Figs. 39–83 View FIGURES 39 – 51. C View FIGURES 52 – 59. C View FIGURES 60 – 65. C View FIGURES 66 – 67. C View FIGURE 68. C View FIGURES 69 – 74. C View FIGURES 75 – 82. C View FIGURE 83. C )
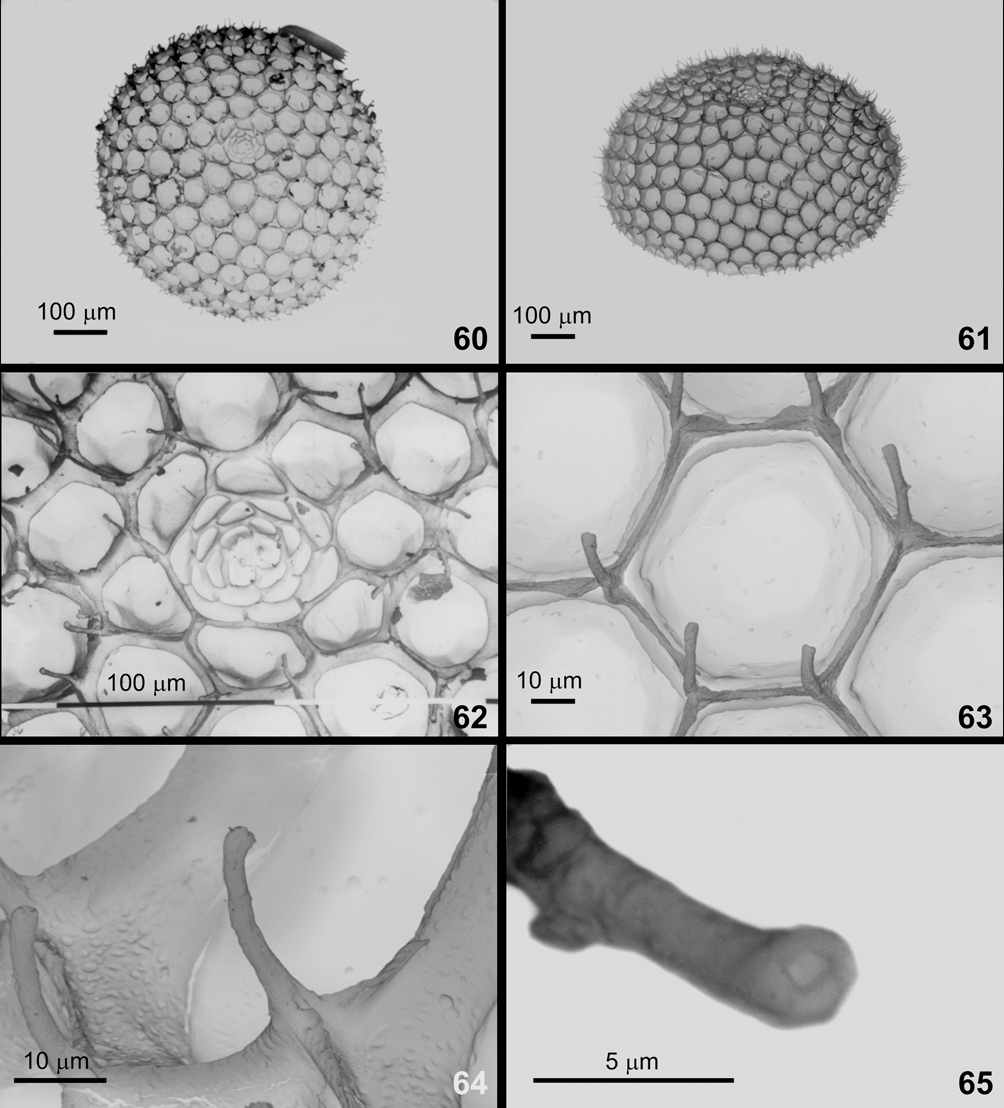
Egg ( Figs. 39, 40 View FIGURES 39 – 51. C , 60–65 View FIGURES 60 – 65. C ). With regard to color pattern, shape, and chorionic ornamentation, the egg of C. janeirica looks like the eggs of C. bellera , C. caulonia , C. cecrops , and C. isobeon . Chorionic “islands” (sensu Downey & Allyn 1981) protrude from floor of rosette cells. There are three micropylar openings in C. janeirica ( Fig. 62 View FIGURES 60 – 65. C ), six in C. caulonia , and three or four in C. bellera .
Diameter 0.72–0.80 mm (X = 0.78 mm, SD = 0.02, N = 35); height 0.40–0.52 mm (X = 0.43 mm, SD = 0.03, N = 35). Duration 6–11 days (X = 9.14 days, SD = 1.01, N = 55).
First instar ( Figs. 41–44 View FIGURES 39 – 51. C , 66–71 View FIGURES 66 – 67. C View FIGURE 68. C View FIGURES 69 – 74. C , 75, 76 View FIGURES 75 – 82. C ). Morphologically similar to that of C. bellera and C. caulonia , but distinguished by the presence of whitish maculae at base of dorsal and lateral setae ( Fig. 42 View FIGURES 39 – 51. C ). Mandibles of first instar of C. janeirica resemble those of C. bellera with six teeth and no tooth on oral surface ( Figs. 69, 70 View FIGURES 69 – 74. C ).
Head capsule width 0.24–0.32 mm (X = 0.28 mm, SD = 0.03, N = 35). Body length ranging from 1.20 mm to 3.52 mm (X = 2.33 mm, SD = 0.85, N = 30). Duration 4–8 days (X = 6.05 days, SD = 0.80, N = 55).
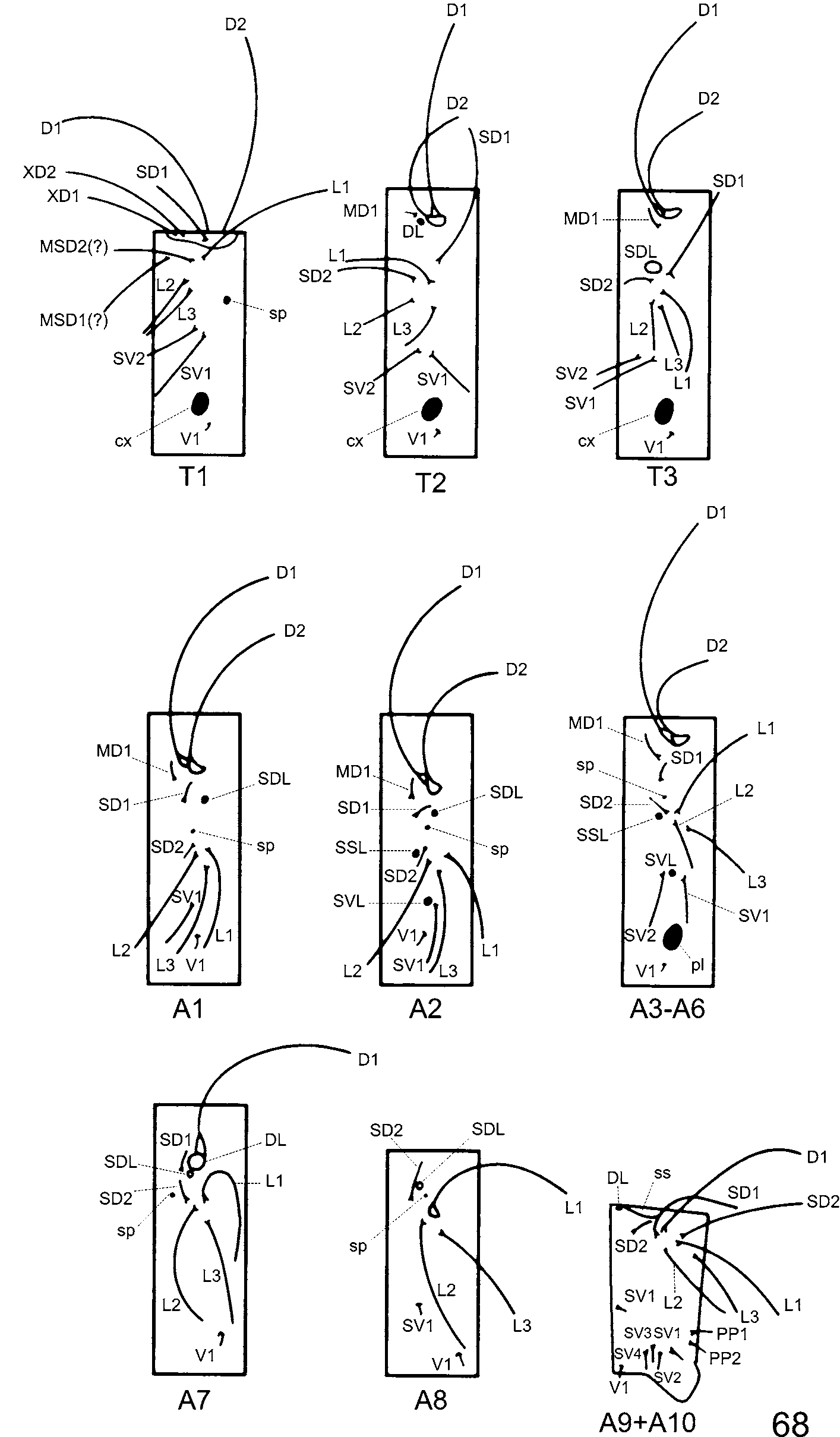
Head chaetotaxy ( Figs. 66, 67 View FIGURES 66 – 67. C ). Same number of primary setae and pores as C. bellera and C. caulonia , and as in C. bellera , setae AF2, CD3, F1, F2, L1, and P2 are absent in C. janeirica . A1, A2, AF1, C1, and C2 usually shorter than those of C. bellera ( Fig. 66 View FIGURES 66 – 67. C ), but size of setae vary intra- and interspecifically ( Duarte et al. 2005).
Labrum chaetotaxy. Identical to that of C. caulonia ( Duarte et al. 2005: 12, fig. 29) and C. bellera .
Body chaetotaxy ( Fig. 68 View FIGURE 68. C ). Same number of primary setae and pore cupola organs as in C. bellera (this paper), distributed as follows:
Prothorax ( Fig. 68 View FIGURE 68. C ). Identical to that of C. bellera and C. caulonia first instar (see Duarte et al. 2005: 13, figs. 30, 32).
Mesothorax ( Fig. 68 View FIGURE 68. C ). Identical to that of C. bellera and differing from C. caulonia in having MD1 much shorter, about a tenth length of D1.
Metathorax ( Fig. 68 View FIGURE 68. C ). Identical to that of C. bellera and C. caulonia first instar (see Duarte et al. 2005: 12, 15, fig. 30), including a conspicuous subdorsal pore cupola organ (SDL) of almost half the width of metathorax in all species.
Abdominal segment 1 ( Fig. 68 View FIGURE 68. C ). As in C. bellera , with 10 pairs of setae and one pair of subdorsal PCOs (SDL of Ballmer & Pratt 1992). D2 slightly shorter than D1. Unlike C. caulonia , subventral PCOs (SVL of Ballmer & Pratt 1992) are lacking in C. janeirica as well as in C. bellera .
Abdominal segment 2 ( Fig. 68 View FIGURE 68. C ). Similar to that of C. bellera and C. caulonia ( Duarte et al. 2005: 15, fig. 30). D1 and D2 as in previous abdominal segment.
Abdominal segments 3–6 ( Fig. 68 View FIGURE 68. C ). Identical to those of C. bellera , but missing a pair of subdorsal PCOs on each segment (SDL of Ballmer & Pratt 1992). Distinguished from those of C. caulonia first instar by absence of V2 ( Duarte et al. 2005: 12, 15, fig. 30).
Abdominal segment 7 ( Fig. 68 View FIGURE 68. C ). As in C. bellera , C. cecrops , C. isobeon and C. caulonia , with a dorsal PCO (DL of Ballmer & Pratt 1992) joined to chalaza of D1. Differs from C. bellera by lacking SV1 and subventral PCOs (SVL of Ballmer & Pratt 1992). These PCOs also absent in first instar of C. caulonia ( Duarte et al. 2005: 12, 15, fig. 30).
Abdominal segment 8 ( Fig. 68 View FIGURE 68. C ). Differs from C. bellera in having L3 almost as long as L2; SV1 much shorter, nearly equal to length of V1 as in C. caulonia . Unlike both C. bellera and C. caulonia , subventral PCOs missing.
Abdominal segments 9+10 ( Fig. 68 View FIGURE 68. C ). Differs from C. bellera by the presence of SD2 on ninth segment. Differs from first instar of other Calycopis with chaetotaxy available by presence of SV1 on A9. Length and position of setae similar to those of C. caulonia .
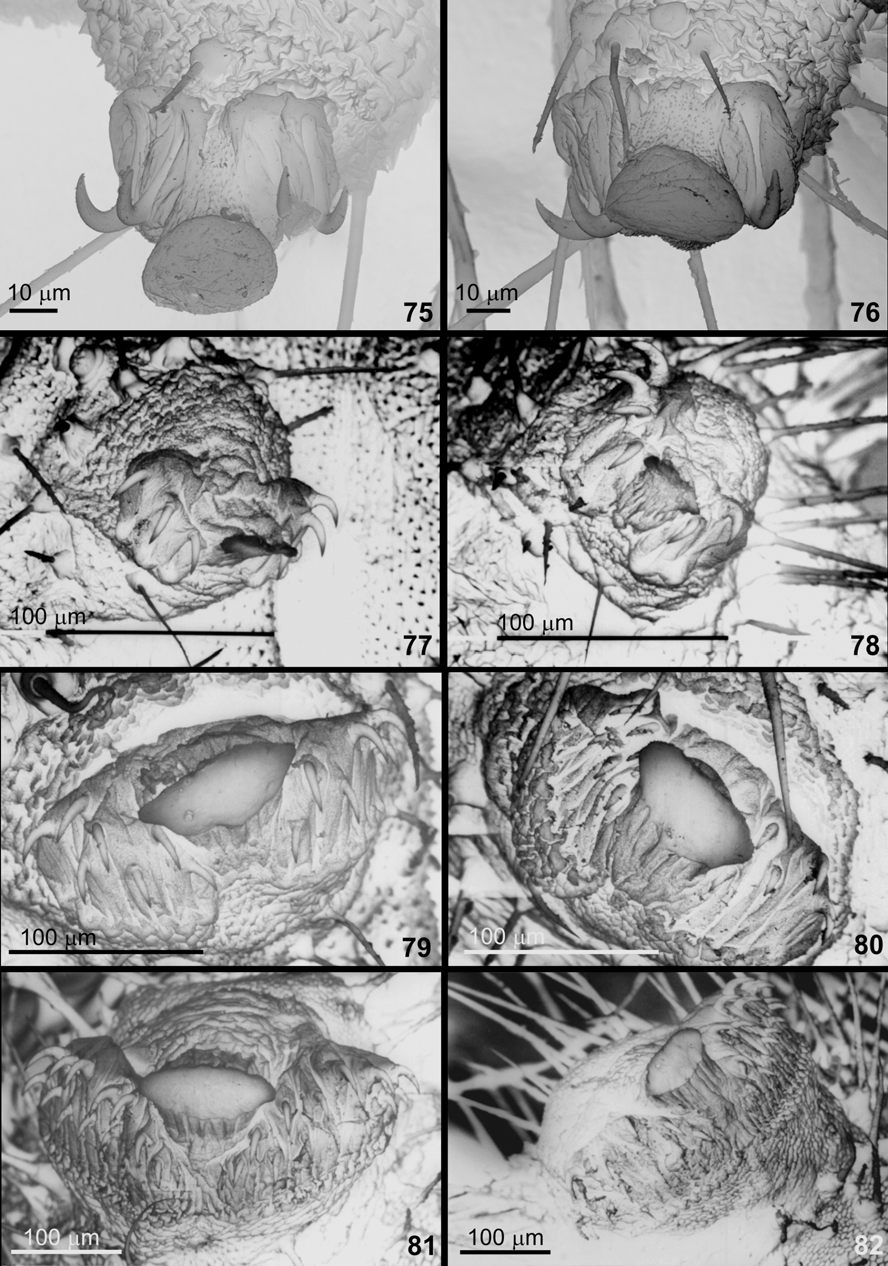
Second instar ( Figs. 45 View FIGURES 39 – 51. C , 72 View FIGURES 69 – 74. C , 77, 78 View FIGURES 75 – 82. C ). Cephalic color pattern resembling that of C. bellera and C. caulonia . Mandibles identical to those of C. bellera with six teeth visible externally as well with a toothlike process located ventrally on oral surface ( Fig. 72 View FIGURES 69 – 74. C ); same number of setae as first instar. Integument coloration identical to C. bellera , including the two longitudinal reddish stripes extending dorsally and laterally from mesothorax to last abdominal segment. Unlike C. bellera , C. caulonia , C. isobeon , and C. vitruvia ( Duarte et al. 2005; Duarte unpubl.), dendritic setae (sensu Ballmer & Pratt 1992) missing on this and later instars. Prothoracic shield pentagonal with number and position of setae and pore cupola organs variable. Other characteristics identical to C. caulonia and C. bellera : prothorax wider and shorter than other thoracic segments; integument highly sculptured and distinctive, with uniformly-spaced oval depressions over epicuticular surface; pattern of oval depression similar in remaining instars; prolegs with uniordinal crochets in uniserial mesoseries, interrupted near center by a conspicuous fleshy pad ( Figs. 77, 78 View FIGURES 75 – 82. C ), separating the crochets into anterior and posterior groups.
Head capsule width 0.38–0.52 mm (X = 0.46 mm, SD = 0.04; N = 30). Body length 3.68–6.16 mm (X = 4.31 mm, SD = 0.64, N = 30). Duration 3–5 days (X = 3.96 days, SD = 0.61, N = 54).
Third instar ( Figs. 46, 47 View FIGURES 39 – 51. C , 79, 80 View FIGURES 75 – 82. C ). Similar to previous instar. Differing in having greater number of spiculate setae on thorax and abdomen, prothoracic shield sagittiform, and prolegs with biordinal crochets in uniserial mesoseries ( Figs. 79, 80 View FIGURES 75 – 82. C ). The typical velvet aspect produced by numerous dendritic setae and found in mature larvae of C. bellera and C. caulonia is lacking in C. janeirica .
Head capsule width 0.62–0.76 mm (X = 0.68 mm, SD = 0.07; N = 30). Body length 7.36–8.16 mm (X = 7.63 mm, SD = 0.24, N = 30). Duration 4–8 days (X = 6.87 days, SD = 1.03, N = 52).
Fourth instar ( Figs. 48–51 View FIGURES 39 – 51. C , 73, 74 View FIGURES 69 – 74. C , 81, 82 View FIGURES 75 – 82. C ). Similar to previous instar in shape and color of head, but with a greater number of secondary setae concentrated on antero-medial region of frontoclypeus. Mandibles with three setae rather than two ( Figs. 73, 74 View FIGURES 69 – 74. C ). Integument orange-yellow with numerous golden secondary setae ( Figs. 48–51 View FIGURES 39 – 51. C ). Prothoracic shield in two symmetrical parts. Prolegs with triordinal crochets in uniserial mesoseries ( Figs. 81, 82 View FIGURES 75 – 82. C ).
Head capsule width 0.92–1.16 mm (X = 1.02 mm, SD = 0.11; N = 30). Body length 8.96–17.60 mm (X = 13.96 mm, SD = 4.13, N = 30). Duration 9–15 days (X = 12.23 days, SD = 1.511, N = 47).
Pupa ( Figs. 52–59 View FIGURES 52 – 59. C , 83 View FIGURE 83. C ). Similar to those of C. bellera and C. caulonia . Head and thorax initially translucent light yellow ( Figs. 52, 53 View FIGURES 52 – 59. C ). Abdomen dorsally with intersegmental pale red maculae extending laterally, but not reaching spiracles ( Figs. 52, 53 View FIGURES 52 – 59. C ). Integument darkening gradually to light brown, with dark brown maculae and small golden prominent setae scattered on dorsal and lateral body ( Figs. 54–57 View FIGURES 52 – 59. C ). Differing from all other reared Calycopis species by the presence of dendritic setae around A6 spiracle ( Fig. 83 View FIGURE 83. C ).
Pupal width on metathorax 2.88–4.96 mm (X = 3.86 mm, SD = 0.54, N = 30), on segment A3 3.68–5.76 mm (X = 4.63 mm, SD = 0.57, N = 30). Pupal length 6.72–11.04 mm (X = 9.33 mm, SD = 1.15, N = 30). Duration 9–14 days (X = 13.00 days, SD = 1.07, N = 46).
Development on artificial diet. Caterpillars of C. janeirica have four instars rather than five as in C. bellera and C. caulonia . All larval instars of C. janeirica fed on artificial diet, which supported complete development. Data on development times from egg to adult are summarized in Table 1 View TABLE 1 . Females of C. janeirica emerged from pupae as adults earlier on average (X = 50.06 days, SD = 2.07, N = 18) than males (X = 52.39 days, SD = 1.71, N = 28), as was the case with C. caulonia and C. bellera .
Cannibalism. As with other Calycopis , when mature larvae were confined in containers, cannibalism was frequent.
Taxonomy. The previous identification of C. janeirica was incorrect ( Field 1967a; Johnson 1988, 1991), and the reasons for our identification of this species were outlined above. In southern Brazil, including our reared adults, the male dorsal hindwing lacks blue and the female dorsal hindwing has blue, but the extent of this coloration in the female is variable. In other parts of South America, the amount of dorsal hindwing blue is variable in both sexes, and the number of species involved is yet to be determined.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |