
Perothopinae, Lacordaire, 1857
|
publication ID |
https://doi.org/ 10.11606/1807-0205/2020.60.special-issue.28 |
|
persistent identifier |
https://treatment.plazi.org/id/038A5C1E-CA12-E06C-3DDC-46D72D32FA2A |
|
treatment provided by |
Carolina |
|
scientific name |
Perothopinae |
| status |
|
Perothopinae , Phyllocerinae , Anischinae?
The larvae of the remaining three basal Eucnemidae subfamilies ( Perothopinae , Phyllocerinae , Anischinae) are very different from each other.
Anischia has microtrichial patches and antennae placed in deep cavities, both features of eucnemids. Its position between Palaeoxenus and ( Melasinae ( Eucneminae , Macraulacinae )) ( Lawrence et al., 2007) appears problematic, as Anischia possesses several plesiomorphic features hard to see as apomorphic reversals needed for such a placement: unmodified head capsule, elongate, normal mandibles and legs with one real segment. We find this result untenable. To place Anischia basally as the sister group of ( Pseudomeninae (Schizophilinae (DE))) is a reasonable possibility. It would mean that the larval labrum was fused to head a least twice in the Eucnemidae lineage, once in Anischia and again in Schizophilus . On the other hand, Perothops with many plesiomorphic features, e.g., antennae attached to surface of head, has a fused larval labrum as well. Thus it is clear that this character state cannot be optimized without multiple changes anyway, irrespective of the cladogram. The free larval labrum in Phyllocerinae and Pseudomenes point to the basal position of eucnemids in the elateroid phylogeny, as the only other taxon to show this plesiomorphic feature is Brachypsectra.
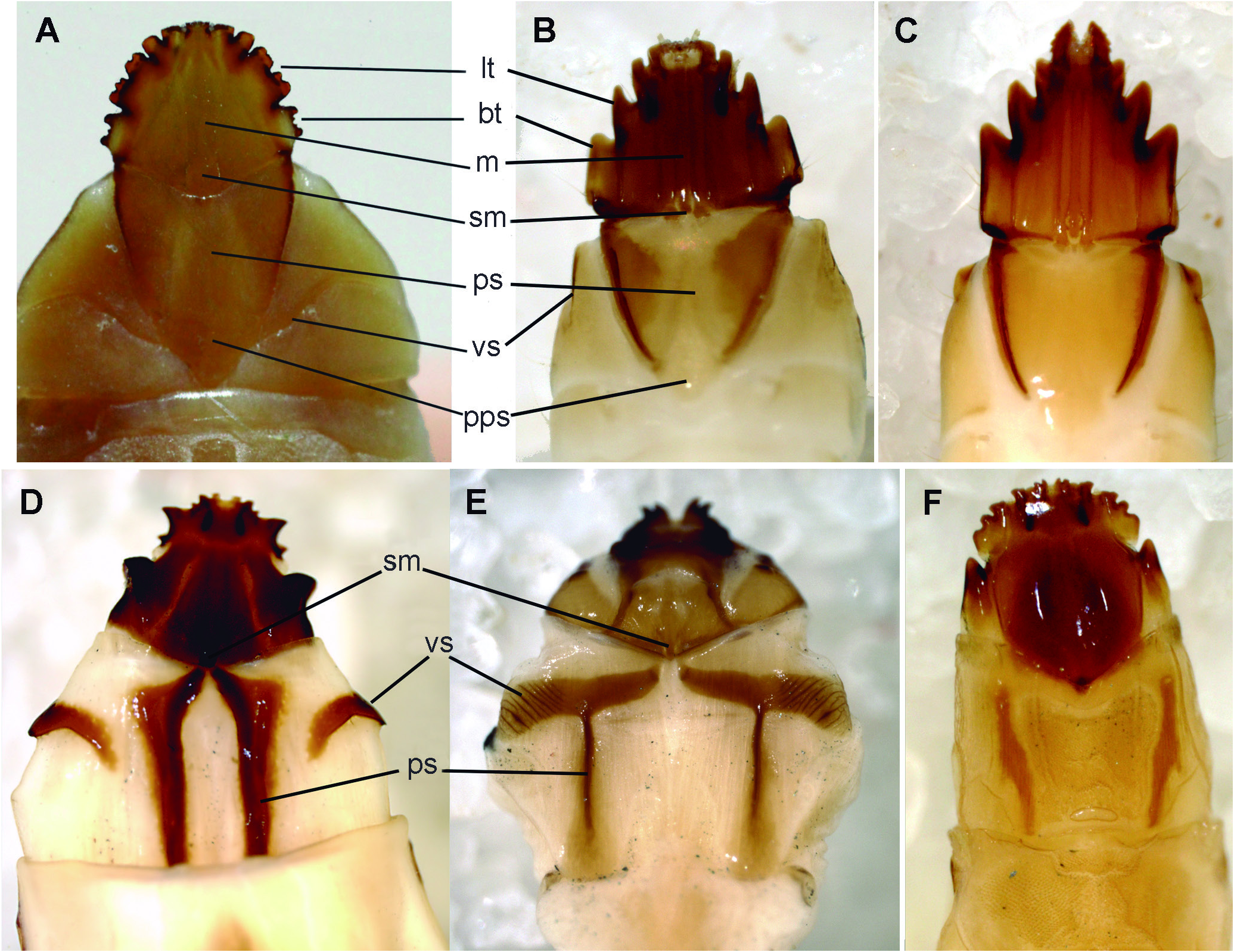
The maxillolabial complex is contiguous with epicranium on each side and incapable of movement in Perothops, Anischia, Pseudomenes and Schizophilus as well as in Cerophytidae and Throscidae , whereas it is solidly fused to epicranium in Phyllocerus as well as in the DE.These fusions are not similar, however. In Phyllocerus , all ventral structures form one single hard plate without details. In Palaeoxenus the separate structures of ventral head as well as prothorax are still discernable ( Fig. 7A View Figure 7 ), whereas in Melasinae , Eucneminae and Macraulacinae the fusion is nearly complete and only mentum and/or submentum can be seen ( Figs. 7 View Figure 7 D-F).
Elateridae as well as all potential out-groups have a free maxillolabial complex, the contiguous state being present in cerophytids, throscids and Perothops, Anischia, Pseudomenes and Schizophilus. This could be a synapomorphy connecting the three clades together. Phyllocerus larva is strongly sclerotized and highly specialized for living in the black earth soil. The apparent similarity in larval maxillolabial complex to lignicolous derived eucnemids is not due to identical structures, but superficial similarity. Phyllocerus larva has deep antennal cavities, however. This might be seen as further support for closer relationship with eucnemids, excluding Perothops .
Thefusedmaxillolabialcomplexappearstobeanother previously ignored synapomorphy for the (Pseudomenini ( Schizophilini (DE))) clade. The sclerotized dark plates and rods found in most Melasinae , Eucneminae and Macraulacinae larvae may be homologs of larval features still visible in Pseudomenes, Schizophilus and Palaeoxenus. The longitudinal rods/fields could be the remnants of the powerful presternal oblique carinae and the paired sclerotization on front corners of proventer homologs of the sclerotized ventral suture edges ( Figs. 7 View Figure 7 D-F).
Do the larval characters support existing hypothesis for Eucnemidae relationships?
Do our undoubtedly speculative ideas (based on actual character states, however) fit the adult structures and the hypothesis based on those? Traditionally eucnemids were separated from elaterids by the completely connate abdominal ventrites and concealed labrum. Neither of these characters suffices today. Anischia fails on both counts with three connate ventrites and a visible labrum. Phyllocerus has only four connate ventrites as do Cerophytidae and Elateridae , whereas Perothops , Pseudomeninae , and all derived Eucnemidae have five connate ventrites,but so does Throscidae . A further character, the attachment of pedicel to scape is subapical in all Eucnemidae , but the same holds for Cerophytidae and a few Elateridae ( Thylacosterninae , Lissominae ). Numerous other characters prove this state in elaterids to be homoplastic, but the cerophytids might well be the sister-group to some or all eucnemids based on the antennal structure.
The female reproductive tract seems to provide illuminating evidence. First, potential out-groups Dascillidae and Artematopidae as well as Throscidae , Cerophytidae and Elateridae all have special sclerites or asperities in bursa copulatrix, Eucnemidae and Brachyspectridae do not (c73). Secondly, Elateridae have colleterial glands, while Eucnemidae , Cerophytidae and Throscidae do not (c70). Third, Perothops , Phyllocerinae , Pseudomeninae , Palaeoxenus and Phlegon Laporte as well as basal Melasinae (e.g., Hylochares Latreille ), Eucneminae (e.g., Phaenocerus Bonvouloir, Pinaroo Muona ) and Macraulacinae (e.g., Langurioscython Heller, Dictyeucnemis Lea, Ceratogonys Perty ) have a bifurcate bursa, a synapomorphy not known in other Elateroidea (c72; Muona, 1993).
The unarmed bursa is clearly a synapomorphy for Eucnemidae s.l. as it is not present in any putative outgroup (c72). The bifurcate bursa is a synapomorphy for eucnemids as well, but it has reversed in all derive members of Macraulacinae , Eucneminae and Melasinae , quite clearly independently in all three clades.
The colleterial glands appear to be the only elaterid synapomorphy found this far, all other related taxa lacking these. It is a strong character not known to have reversed.
Otto & Gruber (2016) drew attention to the peculiar grooves and large punctures in the elytral apices of the eucnemid Stethon . These specialized pores,not previously noticed, had not caught the attention they deserved. A search has shown that these structures (excretory organs?) are found in all Eucnemidae except Perothops, Phyllocerus and a few highly derived Macraulacinae . This splendid discovery appears to be a synapomorphy for all true lignicolous eucnemids.
The hypermetamorphic taxa, i.e., many if not all Eucneminae , many Dirhagini and all Nematodini , are not identical either in structure or in development. There appears to be a pattern present in many Melasinae , Eucneminae and Macraulacinae of early triungulin type larvae followed by chisel-shaped head type stage and finally variable reduction of hard structures in the last stage. Either of the two final stages may dominate the full development.
The extreme larval forms, type Galbitini , resembling Diptera larvae may spend their whole life in one chamber.The extraoral digestion in wood infested with fungus makes this approach feasible.
One of the authors (JM) has followed the development of an undescribed Nematodes Berthold species in Viti Levu, Fiji. The first instar larva had powerful exodont mandibles, similar to the macraulacine Fornax gardneri Fleutiaux first instar larva ( Gardner, 1926). These larvae were found in seemingly hard wood of an unidentified rattan. They developed into a typical chisel-shaped head type in a few weeks and during the same period the substrate had become soft and penetrable. After several such stages the larvae assumed the Nematodes appearance described by Leiler (1976) and Otto (2017). Finally, the further reduced prepupal form emerged. The key here, as with Melasini and Hylochares , seems to be the consistency of the substrate. All these species start their development in hard wood infested by fungus. Early on the larvae need to be able to penetrate the substrate with strong mandibles, but later in their development, if the wood becomes soft enough, a standard derived eucnemid larval type is the solution.
Lignicolous larvae are constantly in danger of being parasitized or eaten by predators. The hydraulic method of penetrating the wood used by eucnemid larvae leaves no real galleries and the absence of working mandibles reduce the noise produced substantially. It is definitely the most advanced approach for moving around within wood in secrecy. The extraoral feeding leaves open the further possibility of not moving at all, if fungal growth is constantly available for extraoral feeding. This may well be the reason behind the extreme larval forms,which appear to spend the whole life in one chamber. It is easy to understand, that such a solution would also benefit from highly mobile early stages capable of finding the right one location, a triungulin type larva.
The few derived eucnemids with larvae penetrating hard wood need strong mandibles and a large prothorax with strong muscles and gripping surface structures to provide the mouthparts leverage. Other beetle larvae with the same kind of approach to wood-boring have developed similar looking solutions, e.g., Buprestidae and Cerambycidae . Details differ substantially; however, those taxa have a free labrum and normally biting mandibles instead of fused labrum and exodont mandibles.
In our view the external similarity of Hylocharini , Melasini and Nematodini is due to repeated invasions into hard wood. In Melasini and Hylocharini the whole development takes place in hard wood. In Nematodini , the larvae use different phases of rot during their development. This may well be the strategy for Eucnemis as well. We have observed minute larvae in moist, very soft inner wall of hollow trees, with larger larvae in more solid and drier wood, deeper in the substrate.
We acknowledge the fact that evolutionary hypotheses must be based on global analytical results. Evolutionary novelties, synapomorphies, exist only in connection with an analysis of a specific data set. On the other hand, we believe it is useful to examine the characters supporting the groups previously analytically established. We have here looked at larval features not used in the evolutionary hypothesis previously produced. The larval features discussed appeared to help to clarify the relationships within Eucnemidae and Elateroidea to some extent. They were especially useful in pointing out serious problems as well as strong evolutionary signals within Eucnemidae .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.