
Tyrannosaurus rex
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2004.00130.x |
|
DOI |
https://doi.org/10.5281/zenodo.3482998 |
|
persistent identifier |
https://treatment.plazi.org/id/03858796-EA19-FF99-8DEA-FE40FBBBF8E6 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
TYRANNOSAURUS REX OSBORN, 1905
Aublysodon molnaris (Leidy, 1868) - Paul, 1988b: p. 325 View Cited Treatment .
Aublysodon cf. A. mirandus (Leidy, 1868) - Molnar & Carpenter, 1989: figs 1-4, p. 447 View Cited Treatment .
Stygivenator molnari (Paul, 1988b) - Olshevsky & Ford, 1995: fig. 36, p. 116 View Cited Treatment .
Albertosaurus cf. A. lancensis Molnar, 1980 : figs 1-7, p. 102 View Cited Treatment .
Albertosaurus megagracilis Paul, 1988b : p. 333. View Cited Treatment
Nanotyrannus lancensis Bakker, Williams, & Currie, 1988: pls. 1-4, figs 1-7, 11, 12, p. 2 View Cited Treatment .
Dinotyrannus megagracilis Olshevsky & Ford, 1995 : fig. 37, p. 117 View Cited Treatment .
Type species: Tyrannosaurus rex Osborn, 1905 .
Discussion and description
The first skeletal material referred to cf. Aublysodon was an isolated frontal ( TMP 80.16.485) from the Dinosaur Park Formation of Alberta ( Currie, 1987). Currie tentatively referred the frontal to Aublysodon because, in contrast to Albertosaurus libratus , the frontal is relatively thin and narrow for a given length. The presence of Aublysodon in the Dinosaur Park Formation was supported by the presence of isolated nondenticulate premaxillary teeth and ‘gracile’ denticulate maxillary and dentary teeth, thought to be referable to the taxon ( Currie, 1987: 55). Currie suggested that the Jordan Theropod might be referable to Aublysodon .
Lehman and Carpenter (1990: 1026; figs 1–6 View Lehman and Carpenter 1990: Figure 1 View Lehman and Carpenter 1990: Figure 2 View Lehman and Carpenter 1990: Figure 3 View Lehman and Carpenter 1990: Figure 4 View Lehman and Carpenter 1990: Figure 5 View Lehman and Carpenter 1990: Figure 6 ) referred a partial skeleton to Aublysodon that is likely referable to a new genus of basal tyrannosauroid. Molnar & Carpenter (1989) described aspects of the specimen, LACM 28471 View Materials , pertinent to a revision of Aublysodon . Their referral of the specimen to Aublysodon cf. A. mirandus was based on the presence of an associated nondenticulate premaxillary tooth. The specimen provided the opportunity to ascribe skeletal characters to the taxon. In their abstract, Molnar & Carpenter (1989: 445) listed one skeletal and one dental character to characterize Aublysodon : a ‘peculiar first dentary tooth’ and a V-shaped frontoparietal suture ( Fig. 2A, B, L View Figure 2 ). Their revised diagnosis of A. mirandus included the following additional characters: slender dentaries, an emargination in the dentary caudal to the third alveolus, and an acute angle between the dorsal and rostroventral margins of the dentary ( Fig. 3D, E View Figure 3 ). They also listed additional dental characters for A. mirandus : the mesial carina of the first dentary tooth is displaced lingually ( Fig. 3E View Figure 3 ), all dentary teeth but the first ( Fig. 3D, E View Figure 3 ) are laterally compressed as well as those of the maxilla ( Figs 3D, E View Figure 3 , 4 View Figure 4 ), denticles extend midway down the mesial carina and to the base on the distal carina and the mesial denticles are smaller than the distal ones.
Figure 1. Continued
Figure 2. Continued
Comparison of the Jordan theropod ( LACM 28471 View Materials ) with tyrannosaurid material weakens the validity of A. mirandus . All tyrannosaurids, except adult Daspletosaurus and Tyrannosaurus , have V-shaped frontoparietal sutures. Slender dentaries are typical of juvenile tyrannosaurids ( Carr, 1999). Regarding the emargination in the dentary, in tyrannosaurids the alveolar margin is convex adjacent to the rostral four or five teeth and is then concave through the midlength of the bone ( Fig. 5 View Figure 5 ). The ‘step’ is the transition point between these margins, and in LACM 23871 View Materials this is exaggerated by the fragmentary nature of both bones ( Figs 3 View Figure 3 D, E, 5). Molnar & Carpenter (1989: 447) cite the ‘rostroventral border of dentaries with an acute angle relative to alveolar border’ as unique. However, this condition is typical of small tyrannosaurid dentaries ( Fig. 5 View Figure 5 ).
Molnar & Carpenter (1989) considered the extent and coarseness of denticles to be diagnostic for A. mirandus . They stated that in the dentary and maxillary teeth of LACM 28471 View Materials , the mesial denticles only extend halfway down their carinae and the entire extent of the distal carinae is denticulate. We find that this character is typical of most tyrannosaurids, except Daspletosaurus torosus in which the mesial carina reaches the crown base, and is therefore not diagnostic of A. mirandus . Also, the condition of mesial denticles being finer than the distal denticles is typical of tyrannosaurids ( Carr & Williamson, 2000) and is not a diagnostic character of any species.
The partial skull of LACM 28471 View Materials displays many characters typical of Tyrannosauridae . The frontal is diagnostic among Late Cretaceous theropods ( Currie, 1987). The bone displays most of the diagnostic tyrannosaurid features, including the following: separate joint surfaces of the lateral and medial frontal processes of the nasal; short orbital rim; expanded frontopostorbital suture with a buttress (although undeveloped in LACM 28471 View Materials ) and caudolateral suture; frontals separated on the midline by the parietals; frontals flat between the orbits; dorsotemporal fossa covering much of the dorsal surface of the frontals ( Fig. 2A- E, G View Figure 2 ).
LACM 28471 View Materials displays additional tyrannosaurid characters, including a first maxillary tooth that is small and incisiform ( Figs 1D, E View Figure 1 , 4 View Figure 4 ) and a subconical first dentary tooth that is the smallest member of the tooth row ( Fig. 3 View Figure 3 D, E). In lateral view, a large neurovascular foramen pierces the maxilla rostral to the antorbital fossa ( Fig. 1D View Figure 1 ), the alveolar margin of the maxilla is convex ( Fig. 1D, E View Figure 1 ) and that of the dentary is concave caudally and convex rostrally ( Fig. 3 View Figure 3 D, E); the rostral-most neurovascular foramen that pierces the lateral surface of the dentary is the largest of its row and is situated ventral to the first alveolus ( Fig. 3 View Figure 3 E). This foramen is also in dromaeosaurids ( Paul, 1988a). In dorsal view, the nasals are constricted at midlength between the maxillae ( Fig. 1A– C View Figure 1 ); the parietals are wedge-shaped and bear a prominent sagittal crest ( Fig. 2I –M View Figure 2 ). Finally, in ventral view, the parietals possess a plug-like process on the rostral midline for apposition with the frontals ( Fig. 2M View Figure 2 ). Thus, there are no characters that contradict assignment of LACM 28471 View Materials to Tyrannosauridae .
Taxonomic identity
As in Daspletosaurus torosus and Tyrannosaurus rex , the lingual surface of each of the third left and right maxillary crowns in LACM 28471 View Materials bear a prominent apicobasal ridge that creates a crease adjacent to the mesial carina ( Fig. 4 View Figure 4 ). In other tyrannosaurids (e.g. Albertosaurus ), this ridge and sulcus are absent. As in juvenile (e.g. CMNH 7541 ) and adult (e.g. AMNH 5027 View Materials ) T. rex , the rugose lateral surface of the maxilla stops far short of the proximal end of the ascending process of the maxilla ( Figs 1F View Figure 1 , 6 View Figure 6 ), indicating that the antorbital fossa reached the nasal suture. Also, the sagittal crest is present on the frontals, a state absent in tyrannosaurids other than Daspletosaurus and Tyrannosaurus .
Several characters of LACM 28471 View Materials are shared with T. rex . As in juvenile T. rex (e.g. CMNH 7541 ), the dorsal and lateral surfaces of the rostral third of the nasals are set at abrupt angles to one another, such that the bones are angular in cross-section ( Fig. 1A View Figure 1 ). This is also found in dromaeosaurids ( Paul, 1988a). The nasals do not separate the distal ends of the nasal processes of the premaxillae by a pair of median ridges, indicating that the processes were appressed through their entire length, as in T. rex (e.g. AMNH 5027 View Materials , CMNH 7541 ; Fig. 7 View Figure 7 ). On the basis of these shared derived characters, we consider LACM 28471 View Materials to be referable to Tyrannosaurus rex .
Ontogenetic stage
In addition to its small size (estimated skull length: ~450.0 mm), LACM 28471 View Materials shares many features that typify juvenile tyrannosaurids (sensu Carr, 1999). In general, the teeth are labiolingually narrow (‘bladelike’) and the cranial remains are lightly built, especially those of the dorsal skull roof such as the nasals, frontals, and parietals ( Figs 1–3 View Figure 1 View Figure 2 View Figure 3 ). LACM 28471 View Materials represents a tyrannosaurid growth stage that precedes that of ‘small Stage 1’ described by Carr (1999) for Albertosaurus libratus .
Molnar (1978: 74) observed that the joint surface for the maxilla on the nasal is smooth and favourably compared this with dromaeosaurids in contrast to the ‘dentate appearance’ of this region in tyrannosaurids. A serrate nasomaxillary suture is characteristic of large adult tyrannosaurids and that of subadult and juvenile specimens is a smooth tongue-in-groove contact ( Carr, 1999). Thus, the smooth state in LACM 28471 View Materials is a juvenile character ( Fig. 1D–G View Figure 1 ).
The frontal process of the nasal is not constricted between the lacrimals in LACM 28471 View Materials ( Fig. 1B, C View Figure 1 ). The same is also true for juvenile Daspletosaurus (e.g. TMP 94.143.1) in contrast to the constricted state in adults (e.g. CMN 8506, TMP 85.62.1). An unconstricted nasal is present in a juvenile individual of T. rex (e.g. CMNH 7541 ) where only the unmodified right lateral frontal process is preserved ( Gilmore, 1946; Bakker et al., 1988; Carr, 1999). In adult T. rex , the process is compressed to a pair of rods between the lacrimals ( Osborn, 1912; Fig. 8 View Figure 8 ).
As in juvenile T. rex (e.g. CMNH 7541 ), the nasals of LACM 28471 View Materials are not rugose, their condition in large adult specimens ( Fig. 1A–E View Figure 1 ). This smooth condition differs from a small juvenile A. libratus (e.g. TMP 86.144.1), in which the nasals are rugose. As in juvenile and adult tyrannosaurids, the internasal suture is open along the rostral and caudal extremities of the bone, but the remainder is closed in external view ( Fig. 1A–C View Figure 1 ). Thus, growth of this bone was not inhibited by closure of this suture and indicates the functional importance of a stable snout for small juvenile tyrannosaurids.
The maxilla, as in juvenile tyrannosaurids ( Carr, 1999), is dorsoventrally shallow and transversely narrow ( Fig. 1A, D–G View Figure 1 ), indicating a long, low snout of delicate construction. The triangular profile of the maxilla noted by Molnar (1978) typifies the proportions of this bone in juveniles and all small theropods and is not taxonomically diagnostic ( Currie & Dong, 2001b; Fig. 6 View Figure 6 ). Molnar observed that the angle of the ventral and rostrodorsal margins converge at a low angle (50°) in LACM 28471 View Materials . In contrast to larger tyrannosaurids, this difference is growth related, juvenile tyrannosaurids having shallower snouts than adults ( Carr, 1999). This is also indicated by the low angle of the margins of the maxilla, which Molnar (1978: 73) also observed in A. libratus . The low angle in LACM 28471 View Materials , in contrast to the higher angle in A. libratus (70°), is undoubtedly related to the fact that LACM 28471 View Materials was the smallest specimen in Molnar’s sample.
The term ‘maxillary flange’, used herein, refers to the convex ala that extends dorsally from the dorsolateral margin of the maxilla, which overlaps the ventrolateral surface of the nasal. The flange represents the lateral half of a deep longitudinal slot in the maxilla that receives a blade-like process from the ventral surface of the nasal. As in juvenile T. rex (e.g. CMNH 7541), the maxillary flange of LACM 28471 View Materials is present but is not as prominent as that of larger specimens ( Figs 1D–G View Figure 1 , 6 View Figure 6 ).
Comparable to juvenile Tyrannosaurus (e.g. CMNH 7541 , PIN 552–2) and Daspletosaurus (e.g. TMP 94.143.1), the rostral margin of the maxillary fenestra in LACM 28471 View Materials does not approach the rostral margin of the external antorbital fenestra as it does in adults ( Figs 1D, E View Figure 1 , 6 View Figure 6 ). In adult T. rex , the maxillary fenestra extends medial to the rostral margin of the external antorbital fenestra ( Fig. 6 View Figure 6 ), while in Daspletosaurus adults, a T. bataar juvenile (GIN 100/177), and the holotype of Shanshanosaurus huoyanshanensis ( Currie & Dong, 2001a) , the maxillary fenestra is separated from the margin of the external antorbital fenestra by a narrow rim of bone ( Fig. 6 View Figure 6 ). Also as in juvenile tyrannosaurids, in LACM 28471 View Materials a low ridge encircles the rostroventral margin of the antorbital fossa ( Fig. 1D View Figure 1 ), while the ventral foramen within the fossa of the subnarial foramen opens rostroventrally ( Carr, 1999; Fig. 1D View Figure 1 ); in subadults and adults, this foramen is large and opens rostrally ( Carr, 1999).
The dorsotemporal fossa of the frontal is present in LACM 28471 View Materials and, as in juvenile tyrannosaurids ( Carr, 1999), the rostral margin of the depression is indistinct (e.g. CMNH 7541 ; Fig. 2A, B View Figure 2 ) whereas the rostral margin of the dorsotemporal fossa is distinct in large and mature specimens ( Fig. 8 View Figure 8 ). As in other juvenile tyrannosaurids (e.g. CMNH 7541 ), most of the rostral margin of the fossa is not delimited by a distinct ridge and sulcus, except in LACM 28471 View Materials where these features are present at the lateral extent of the fossa in both bones ( Fig. 2A, B View Figure 2 ). Also, the sutural surface of the postorbital is not modified into a dorsoventrally deep abutting and peg-in-socket contact as in larger specimens, but is composed of deep grooves and ridges ( Fig. 2E, G, H View Figure 2 ).
In dorsal view, the joint surface of the lacrimal is rostrocaudally elongate and transversely narrow, as in juvenile tyrannosaurids ( Carr, 1999; Fig. 2A, B View Figure 2 ). This is unlike adults of Daspletosaurus and Tyrannosaurus in which the frontal, caudal to the lacrimal, is transversely wide ( Fig. 8 View Figure 8 ). The same is also true for the prefrontal- frontal suture in T. rex ( Fig. 8 View Figure 8 ). In LACM 28471 View Materials , the joint surfaces of the medial and lateral frontal process of the nasal are separate in dorsal view ( Fig. 2A, B View Figure 2 ), as in other tyrannosaurids. In adult T. rex (e.g. AMNH 5027 View Materials , BHI 3033 , BHI 4100 , FMNH PR 2081 ), the medial frontal process of the nasal is displaced ventrally out of view and the lateral frontal process itself is compressed toward the midline beneath the lacrimal ( Fig. 8 View Figure 8 ). Thus in adults, the joint surface of the medial frontal process is pinched out below and between the compressed nasal processes of the frontal.
As in other juvenile tyrannosaurids ( Carr, 1999), the muscle attachment area on the lateral surface of the surangular, ventral to the glenoid fossa, is a shallow groove ( Fig. 3A View Figure ). In larger and more mature individuals this is a dorsoventrally deep and rugose scar. The shallow dentary of LACM 28471 View Materials also indicates the juvenile status of the specimen ( Figs 3D, E View Figure , 5 View Figure 5 ). In tyrannosaurid growth, dentary depth exhibits positive allometry; in the smallest juveniles, the width of the bone approaches its height.
Molnar (1978) claimed that the dentary of LACM 28471 View Materials is unique with regard to the shallow angle of the symphysis and the slope of the alveolar margin in lateral view. In our view, the low angle of the ventral margin of the symphyseal region relative to the ventral margin of the bone is typical of juvenile tyrannosaurids ( Figs 3D, E View Figure , 5 View Figure 5 ), as noted by Molnar (1978) for A. libratus and Currie & Dong (2001a) for Shanshanosaurus . A steep rostroventral margin of the dentary typifies larger and deeper-jawed specimens. In tyrannosaurids, the alveolar margin of the dentary, in lateral view, is generally concave and becomes convex lateral to the rostral four alveoli ( Fig. 5 View Figure 5 ). This is also the case in LACM 28471 View Materials ( Figs 3D, E View Figure , 5 View Figure 5 ). Therefore, on the basis of these features, LACM 28471 View Materials exemplifies a juvenile T. rex .
REVISED DESCRIPTION
We differ from previous workers in our interpretation of some aspects of the osteology of LACM 28471 View Materials ( Table 1 View Table 1 ). However, we have discovered several details that supplement the previous descriptions ( Molnar, 1978; Paul, 1988b; Molnar & Carpenter, 1989; Olshevsky & Ford, 1995). We were able to verify that the shallow concavity reported by Molnar (1978) along the rostrodorsal margin of the left maxilla is the fossa of the subnarial foramen ( Fig. 1D View Figure 1 ). The ventral margin of the naris would have been delimited by the maxillary processes of the premaxilla and nasal, which are missing in this specimen.
The transverse curvature of the nasals indicates that they were deep and vaulted rostrally ( Fig. 1A, D, E View Figure 1 ). The caudal third of the bone, reported here for the first time, is a flat plate ( Fig. 1B, C View Figure 1 ). Thus, the conjoined bones are constricted at midlength and expand rostrally toward the maxillary processes ( Fig. 1A–C View Figure 1 ). In lateral view, the joint surface for the maxilla extends dorsolaterally behind the maxillary process ( Fig. 1E View Figure 1 ). All of these features are present in other tyrannosaurids. Toward the midlength of the unit, the nasals are inclined dorsally toward the midline, forming a low sagittal ridge along the internasal suture ( Fig. 1A View Figure 1 ), as in juvenile (e.g. CMNH 7541 ) and adult (e.g. AMNH 5027 View Materials ) T. rex . A raised internasal suture is present in other tyrannosaurids (e.g. A. libratus , T. bataar ), but this extends further rostrally in T. rex .
The rostral margin of the antorbital fossa of the maxilla is preserved and it shows the conventional tyrannosaurid contours of a rostrodorsally orientated ventral margin that abruptly extends caudodorsally to form the rostral margin of the fossa ( Fig. 1D View Figure 1 ). This pattern is retained in other juveniles ( Currie & Dong, 2001b; fig. 1). A portion of the ascending process for the maxilla is preserved ( Fig. 1F, G View Figure 1 ). The dorsolateral surface of the bone bears the joint surface for the nasal that distally transfers onto the medial surface deep to the maxillary flange. The medial surface of the process is incised by the dorsoventrally deep joint surface for the lacrimal that tapers rostrally and is situated above the ventral margin of the bone ( Fig. 1G View Figure 1 ).
In LACM 28471 View Materials , the caudolateral margin of the right frontal is present ( Fig. 2B View Figure 2 ) and between the left and right bones the entire joint surface for the postorbital is preserved ( Fig. 2C–E, G View Figure 2 ). The postorbital overlaps the frontal as in other tyrannosaurids ( Fig. 2B–E, G View Figure 2 ). The joint surface for the postorbital consists of two principal components: a rostral buttress and a caudal shelf ( Fig. 2G View Figure 2 ). In lateral view, the rostral buttress is represented by a dorsoventrally deep and transversely shallow cleft. Low ridges that trend caudoventrally to rostrodorsally reinforce this surface. In larger specimens, this surface is dorsoventrally deep and flat, forming an abutting surface that faces caudolaterally and is floored ventrally by a narrow shelf. In LACM 28471 View Materials , this cleft faces laterally ( Fig. 2G, H View Figure 2 ). The floor of the rostral half of the surface continues caudally, and is incised by a deep slot that is overlapped by the caudal shelf. Thus, the caudal shelf fits into a slot in the postorbital ( Fig. 2G View Figure 2 ).
In dorsal view, the caudal half of the joint surface for the postorbital in LACM 28471 View Materials is distinct ( Fig. 2B View Figure 2 ). A slot incises the lateral surface of the bone, whose floor extends laterally in dorsal view ( Fig. 2B–E View Figure 2 ). The slot receives a medial process from the postorbital that extends caudally. In larger specimens, the slot is modified into a series of sockets to receive peg-like processes from the postorbital. In LACM 28471 View Materials , the slot is concealed in dorsal view by a laterally extending shelf ( Fig. 2B–E View Figure 2 ). The dorsal surface of the shelf bears the joint surface, indicating that the structure is clasped dorsally and ventrally by the postorbital. As in other tyrannosaurids, the joint surface is inset relative to the dorsal surface of the bone to abut the medial edge of the portion of the postorbital that overlaps the frontal ( Fig. 2B–E View Figure 2 ). Finally, unlike larger tyrannosaurid specimens (e.g. ROM 1247) in which the postorbital underlaps the caudoventral surface of the frontal, the joint surface of the parietal extends to the lateral edge of the caudal surface of the frontal ( Fig. 2C View Figure 2 ); this is also found in CMNH 7541 .
The dorsal surface of the frontals is not perfectly flat, but concave in the region of the dorsotemporal fossa and flattened at the base of the nasal process, becoming concave again toward the joint surface for the nasal ( Fig. 2A, B View Figure 2 ). This is also found in juvenile T. rex (e.g. CMNH 7541 ).
The frontoparietal suture is serrate laterally ( Fig. 2C, J View Figure 2 ) and medially the suture consists of interleaving grooves and ridges that lie in the horizontal plane ( Fig. 2A, B, I–M View Figure 2 ). Laterally, the bones are in secure contact via stout vertical grooves and ridges ( Fig. 2C, I–M View Figure 2 ). In caudal view, the caudal edge of the frontal overlaps the parietal laterally ( Fig. 2C, I, L View Figure 2 ). Thus, in dorsal view, the parietal overlaps the frontal medially and the frontal overlaps the parietal laterally ( Fig. 2L View Figure 2 ). This pattern is typical of tyrannosaurids.
In lateral view, the joint surface for the prefrontal is the shallow slot that incises the lateroventral surface of the frontal ( Molnar, 1978) and is situated rostroventral to the joint surface of the lacrimal ( Fig. 2F, G View Figure 2 ). Thus, as in other tyrannosaurids, the joint surface is not dorsolaterally directed ( Fig. 2G View Figure 2 ). In ventral view, this surface extends rostral to the joint surface for the lacrimal and widens laterally, allowing the prefrontal to emerge onto the dorsal skull roof medial to the lacrimal and lateral to the frontal ( Fig. 2A, B, F, G View Figure 2 ). Proximally, the joint surface for the prefrontal is shallow, reflecting the absence of a deep socket noted by Currie (1987: 54) that typifies ‘progressive’ tyrannosaurids.
In ventral view, the joint surface for the laterosphenoid on the parietal covers the ventrolateral surface of the bone ( Fig. 2I View Figure 2 ), which in LACM 28471 View Materials consists of a series of coarse slots and grooves that trend rostrolaterally ( Fig. 2I, M View Figure 2 ). The joint surface for the prootic begins caudal to the constricted midregion of the parietal and is represented by longitudinal grooves and ridges. The ventral surface of the parietal fragment is biconcave, with a low transverse ridge that presumably delimits the region of the cerebral hemispheres from that of the midbrain ( Fig. 2M View Figure 2 ).
A fragment of the medial portion of the nuchal crest is present, which includes the sagittal crest ( Fig. 2N- P View Figure 2 ). The sagittal crest is thin and bladelike and reaches the dorsal margin of the nuchal crest, as in CMNH 7541 ( Fig. 2N, O View Figure 2 ). The caudal surface of the fragment retains the thin vertical ridge that widens ventrally to contact the supraoccipital ( Fig. 2O, P View Figure 2 ). The sagittal crest and caudal ridge are offset by a few mm ( Fig. 2O View Figure 2 ); i.e. they are not directly opposite each other. The dorsal surface of the nuchal crest is smooth, indicating the immature status or small size of the specimen ( Fig. 2O View Figure 2 ). The fragment is not complete enough to obtain a contact with the rest of the parietal; it is likely the nuchal crest was transversely orientated, as in juvenile T. rex (e.g. CMNH 7541 ).
Molnar (1978) favourably compared the abrupt constriction between the cerebral and olfactory regions of an endocast of LACM 28471 View Materials with Ceratosaurus and observed that the same passage in T. bataar and T. rex are ‘less abrupt’ (1978: 77). The state of this feature in LACM 28471 View Materials may simply be attributable to immaturity, but a thorough study of a growth series of tyrannosaurid endocasts is required to falsify this inference.
As reported by Molnar (1978), the glenoid region of the right surangular is present ( Fig. 3A–C View Figure ). In dorsal view, caudal to the glenoid, the proximal portion of the joint surface for the articular is preserved and is strengthened by coarse ridges ( Fig. 3B View Figure ). The rostral half of this surface is covered by adherent cortical bone of the articular, indicating that this contact was a secure union.
The Meckelian groove of LACM 28471 View Materials divides the medial surface of the dentary into lingual and ventral bars. Although shallow, the groove is traceable and terminates rostrally at a rostrocaudally elliptical foramen at the level of the fifth alveolus. This is typical of all tyrannosaurids. The bases of three small interdental plates are preserved in the left dentary and six are present on the right bone.
We found that the tooth ‘assumed to be a premaxillary tooth’ ( Molnar,1978: 77) does not simply adhere to the maxilla, but is situated directly opposite the first alveolus, from which it is displaced lingually by several mm ( Fig. 1D View Figure 1 ). This displacement occurs in the second tooth, but to a lesser extreme. There is no bone interposed between the incisiform tooth and the alveolus, precluding the possibility that the tooth was not originally situated in the first alveolus. These observations indicate this tooth is the first maxillary tooth. Although the tooth is incisiform, it has features that are transitional between premaxillary teeth and blade-like maxillary teeth: the distal carina is situated further distally than the lingually positioned mesial carina, and the apical region of the tooth is mesiodistally flattened, such that this region of the tooth is wider than long in cross section ( Fig. 4 View Figure 4 ). This considered, there are six teeth situated rostral to the antorbital fossa ( Fig. 1D View Figure 1 ).
We therefore differ in our opinion from Molnar & Carpenter (1989) of some details of the revised diagnosis and account of Aublysodon mirandus , based on LACM 28471 View Materials . The distolabially shifted carina of the ‘more lateral premaxillary teeth’ actually typifies the first maxillary tooth (1989: 447) and the ‘twist’ in the crown is standard for incisiform teeth in tyrannosaurids in which the apex of the crown deviates labially. This is a useful feature for identifying which side of the skull an isolated incisiform tooth initially came from. One character thought to set Aublysodon apart from tyrannosaurids is a low ridge on the lingual surface of incisiform teeth ( Molnar & Carpenter, 1989); this character is actually typical of tyrannosaurids, including cf. Albertosaurus sp. (e.g. ROM 1422).
We agree with Molnar (1978) that the small first dentary tooth of LACM 28471 View Materials is similar to the symphyseal premaxillary teeth and first dentary tooth of dromaeosaurids. In addition, like dromaeosaurids, the first dentary tooth of tyrannosaurids is the smallest of the tooth row and is conical in form with a linguomesially positioned mesial carina, as in LACM 28471 View Materials ( Molnar & Carpenter, 1989; Fig. 3D, E View Figure ) and the succeeding dentary teeth have the typical laterally compressed form. Also, with few exceptions, mesial denticles in tyrannosaurids tend to extend to the midheight of the crown whereas the denticles of the distal carina always reach the crown base. Smaller mesial than distal denticles is typical of small tyrannosaurid teeth ( Carr & Williamson, 2000). Thus, the ‘diagnostic’ characters of Aublysodon mirandus (sensu Molnar & Carpenter, 1989) are either typical of tyrannosaurids in general or of juveniles in particular.
OTHER PREVIOUS TAXONOMIC WORK ON LACM 28471 View Materials
Paul (1988b) designated LACM 28471 View Materials as the type specimen of the species Aublysodon molnaris . He referred the specimen to Aublysodon based on the low nasals and triangular profile of the maxilla and nondenticulate front teeth. As noted above, the nasals are actually dorsoventrally crushed. The nondenticulate condition of the front teeth is not distinctive and appears to reflect immaturity, as in other theropods such as Velociraptor (e.g. IGM 100/972; Norell et al., 1994). Paul (1988b: 324) created a new species for the specimen based on its larger size, larger teeth, and ‘more robust snout’ in contrast to Shanshanosaurus huoyanshanensis Dong, 1977 (which he referred to Aublysodon ) and A. mirandus . However, these features are vague and hence are not diagnostic. Based on the diagnostic characters given above, we consider A. molnaris to be a junior subjective synonym of T. rex .
Olshevsky & Ford (1995) referred LACM 28471 View Materials to a new genus, Stygivenator molnari . They considered the specimen unique on the basis of its long maxillary teeth and mesiodistally narrow and smaller ‘premaxillary’ tooth, in contrast to the type tooth of A. mirandus . The authors considered the convex rostral end of the alveolar margin of the dentary to be unique; as shown above, this typifies juvenile and small tyrannosaurids ( Fig. 5 View Figure 5 ). Also, they considered the procumbent nature of the rostral three dentary teeth to be diagnostic. However, this can be observed in juvenile and subadult specimens of other tyrannosaurids such as Albertosaurus libratus (e.g. ROM 1247). In our opinion, S. molnari is a junior subjective synonym of T. rex .
LACM 23845 View Materials : DISCUSSION
Taxonomic identity
Molnar (1980) described the partial skull and skeleton ( LACM 23845 ) of a medium-sized tyrannosaurid recovered from a quarry adjacent to one containing an adult T. rex ( LACM 23844 View Materials ) on the L. D. Engdahl Ranch in Garfield County, Montana. He referred the specimen to Albertosaurus cf. A. (= Nanotyrannus ) lancensis . This specimen was later made the holotype of a new species, Albertosaurus megagracilis by Paul (1988b). Olshevsky & Ford (1995) placed this species in a new genus, Dinotyrannus . We suggest that LACM 23845 View Materials represents a subadult T. rex .
LACM 23845 View Materials displays several characters that are shared by Daspletosaurus and Tyrannosaurus . In dorsal view, the external surface of the frontal process of the nasals is constricted between the lacrimals ( Figs 8 View Figure 8 , 9B, D View Figure 9 ). Also, between the constriction and the rostral extent of the joint surface for the lacrimal, the dorsolateral margin of the nasal is notched to receive a stout process from the dorsomedial edge of the lacrimal ( Figs 8 View Figure 8 , 9B View Figure 9 ). In dorsal view, the frontals are transversely wide such that the frontolacrimal and frontoprefrontal sutures are also widened ( Fig. 8 View Figure 8 ). As in subadult and adult specimens of Tyrannosaurus and Daspletosaurus , the sagittal crest is divided on the frontals such that a deep midline cleft separates the paired crests ( Fig. 8 View Figure 8 ). In other tyrannosaurids and in juvenile T. rex (e.g. CMNH 7541 , LACM 28471 View Materials ), a large foramen pierces the sagittal crest on the midline, and the crest diminishes rostrolateral or rostroventral to the opening. The dorsotemporal fossa is deep, and the dorsal surface rostral to the fossa is rostrocaudally short, only 2 cm longer than in the juvenile LACM 28471 View Materials ( Fig. 8 View Figure 8 ). Also, the frontoparietal suture is transversely orientated ( Figs 8 View Figure 8 , 15C, H View Figure ), except on the midline, as in Daspletosaurus and Tyrannosaurus ( Fig. 8 View Figure 8 ). Finally, the lingual ridge and groove are present in an associated crown that is probably a mesial maxillary tooth.
LACM 23845 View Materials displays several Tyrannosaurus characters. As in most specimens of T. rex (except on the left side in LACM 23844 View Materials ), the caudolateral process of the nasal is long ( Figs 9 View Figure 9 , 10 View Figure ). This process is short or absent in Daspletosaurus (e.g. CMN 8506, TMP 85.65.1) and T. bataar (e.g. PIN 553–1), and long in Albertosaurus ( Fig. 10 View Figure ). The lacrimal of LACM 23845 View Materials does not have a cornual process; instead, the dorsal surface is wide and flat, and the accessory pneumatic fossa is located distal to the lacrimal recess ( Figs 10 View Figure , 11C View Figure 11 ). In Daspletosaurus the cornual process is a low mound; in Albertosaurus it is sharply defined, with the dorsal surface of the bone sloping dorsolaterally to the cornual process and the accessory pneumatic fossa proximal in position ( Fig. 10 View Figure ). As in subadult and adult specimens of Tyrannosaurus , the sagittal crest extends rostrally on the frontals such that a midline cleft separates the paired crests ( Fig. 8 View Figure 8 ). In other tyrannosaurids and in juvenile T. rex (e.g. CMNH 7541 , LACM 28471 View Materials ), a large foramen pierces the sagittal crest on the midline and the crest diminishes rostrolateral or rostroventral to the opening.
As in Tyrannosaurus and Albertosaurus , the antorbital fossa depresses the lateral surface of the rostral ramus ahead of the lacrimal pneumatic recess ( Fig. 10 View Figure ). In Daspletosaurus the fossa faces ventrally in large specimens ( Fig. 10 View Figure ). The gentle twist in the rostrolateral margin of the squamosal process of the quadratojugal ( Figs 11G, I View Figure 11 , 12 View Figure ) is less well developed than the notch in the same region of large adult specimens (e.g. AMNH 5027 View Materials , BHI 3033 , FMNH PR 2081 , SDSM 12047 ). This character is diagnostic for T. rex .
In dorsal view, the joint surfaces of the nasal processes of the premaxillae are not separated proximally ( Fig. 9B View Figure 9 ). Although the midline struts that separate the premaxillae on the midline are present, they are set below the external surface of the bone, indicating that the processes were probably appressed throughout their entire length, as in juvenile (e.g. CMNH 7541 ) and adult (e.g. AMNH 5027 View Materials , FMNH PR 2081 ) T. rex ( Fig. 7 View Figure 7 ). As in other specimens of T. rex (e.g. FMNH PR 2081 , LACM 23844 View Materials ), the caudal surangular foramen is smaller than it is in other tyrannosaurids ( Fig. 13 View Figure 13 ). This is also present in some Albertosaurus libratus (e.g. ROM 1247; Fig. 13 View Figure 13 ).
Figure 9. Continued
Finally, two postcranial characters are consistent with other T. rex specimens. In the scapula in ventral view, the caudal margin of the joint surface of the humerus is narrower than the cranial margin (see e.g. FMNH PR 2081 ; Fig. 14C View Figure 14 ), while in lateral view, the acromion is elongate (e.g. FMNH PR 2081 ; Fig. 14A, B View Figure 14 ), characters first noted by Carpenter & Smith (2001). Thus, we consider LACM 23845 View Materials to be referable to T. rex .
Ontogenetic stage
Molnar (1980) provided four reasons why LACM 23845 View Materials is not an immature T. rex , each of which is contestable: presence of a tall nuchal crest, smooth prefrontofrontal and quadratojugoquadrate sutures, and the form of the proximal fibula. The height of the nuchal crest is thought to be ontogenetically variable in tyrannosaurids ( Russell, 1970). This idea is based on the subadult A. libratus specimen AMNH 5664 in which the nuchal crest is ‘only about one-fourth as large as in adults’ ( Russell, 1970: 7). However, the nuchal crest in AMNH 5664 is completely broken off and missing. Prominent nuchal crests are present in much smaller juvenile tyrannosaurid specimens, including T. rex (e.g. CMNH 7541 ; Carr, 1999) and so were presumably originally tall in AMNH 5664. Therefore, the presence of a tall nuchal crest in LACM 23845 View Materials does not indicate adulthood.
In rostral and lateral views, we observed that the central region of the joint surface of the prefrontal in the frontal of LACM 23845 View Materials is coarsened by low papillae and rostrodorsally trending ridges, indicating that the prefrontal- frontal union is not smooth. Thus in T. rex a socket for the prefrontal in the frontal is present late in maturity (cf. Currie, 1987). The shallow and smooth joint surfaces of the quadrate on the quadratojugal ( Molnar, 1980; Fig. 11H, L View Figure 11 ) are interpreted here as size-dependent features in LACM 23845 View Materials . In large adult T. rex the joint surfaces are coarse and deeply excavated (e.g. LACM 23844 View Materials ).
Molnar suggested that the form of the proximal fibula was different from that of Tyrannosaurus . In LACM 23845 View Materials , in lateral view, the proximal end of the fibula is more symmetrical than in Tyrannosaurus , the bone is more slender, and the dorsal margin is flatter ( Molnar, 1980). Although described as symmetrical, the proximal portion of the fibula in lateral view has a nearly vertical rostral margin and the caudal margin dilates proximally. This condition is present in adult T. rex specimens (e.g. FMNH PR 2081 ) and in subadult Albertosaurus (e.g. AMNH 5664, ROM 1247) and so does not appear to reflect relative maturity or size. The slenderness of the bone probably reflects the smaller size of the animal. Molnar (1980: 107) also stated that the ‘projection of the proximal face of metatarsal III’ indicated the specimen’s maturity (Fig. 20H, I, K), but the meaning of this character is unclear.
In our view, LACM 23845 View Materials is probably a subadult T. rex . In addition to a low estimated skull length (~ 80 cm), several features indicate that it was a subadult or small adult at death. Molnar observed that the nasals are not as rugose as in T. rex . However, his comparative material for the latter taxon (e.g. AMNH 5027 View Materials , LACM 23844 View Materials ) consists of larger and more mature specimens; thus the more rugose condition might reflect greater maturity.
As noted by Molnar (1980), the nasal contact of the frontal is W-shaped, but this is due to the subadult status of the specimen. The condition in LACM 23845 View Materials is therefore transitional between the unconstricted condition in juveniles (e.g. CMNH 7541 , LACM 28471 View Materials ) and the constricted condition of adults (e.g. AMNH 5027 View Materials , BHI 3033 , FMNH PR 2081 ; Fig. 8 View Figure 8 ).
The dorsal surface of the lacrimal is situated relatively close to the dorsal margin of the lacrimal recess, indicating that the bone is not as inflated as in large adult specimens (e.g. AMNH 5027 View Materials , BHI 3033 , FMNH PR 2081 , SDSM 12047 ; Carr, 1999; Figs 10 View Figure , 11C View Figure 11 ). The medial joint surface for the nasal on the frontal is visible in dorsal view ( Fig. 15C, D View Figure ). In large adult specimens (e.g. AMNH 5027 View Materials , BHI 3033 , FMNH PR 2081 ), the joint surface for the medial frontal process of the nasal is displaced ventrally out of view between the compressed rods of the frontal process.
Unlike large adult specimens of T. rex , the parasagittal crest of the frontal in LACM 23845 View Materials does not extend far rostrally, but diminishes after a short distance ( Figs 8 View Figure 8 , 15C, D View Figure ). This state is also present in Albertosaurus of any size. In dorsal view, the suture for the prefrontal on the frontal is angular on the left and arcuate on the right in LACM 23845 View Materials ( Fig. 15C, D View Figure ). The asymmetrically developed dorsal surface of the nuchal crest is not developed into a rugose platform as in large adult T. rex (e.g. AMNH 5027 View Materials ), and so might indicate immaturity ( Fig. 15J- M View Figure ). In LACM 23845 View Materials , only the right half of the crest is thickened, but with a smooth dorsal surface ( Fig. 15J- M View Figure ).
The tubercle for attachment of the tendon of M. depressor mandibulae of the otoccipital ( Fig. 16A, B View Figure 16 ) is low, and not prominent as in large adults (e.g. AMNH 5027 View Materials , FMNH PR 2081 ). The margin of the external mandibular foramen of the angular is developed into a rugose ridge ( Fig. 17C View Figure 17 ). A low irregular scar is present in juvenile tyrannosaurids ( Carr, 1999). The prearticular of LACM 23845 View Materials is typical of subadult tyrannosaurids: the caudal ramus is dorsoventrally deep, the dorsal and ventral margins converge toward each other rostrally, and the bone is dorsoventrally flattened and transversely wide at midshaft ( Fig. 17D–G View Figure 17 ). Unlike adult specimen LACM 23844 View Materials , the medial surface of the bone is not excavated by the joint surface for the splenial, the bone is shallower, the rostroventral surface of the joint surface for the angular is flat and not concave, and the bone is narrower at midshaft. As in mature tyrannosaurids, the dentary is much deeper than wide ( Fig. 18A, B View Figure 18 ). Thus, LACM 23845 View Materials displays features that are intermediate in development between juveniles (e.g. CMNH 7541 ) and adults and therefore may be a subadult or small adult T. rex .
Figure 11. Continued
Figure 14. Continued
Figure 15. Continued
Revised description
Our opinion of some details of the osteology of LACM 23845 View Materials differs from that of Molnar (1980). We summarize the differences in interpretation in Table 2 View Table 2 , but several of his observations require comment here.
We found that the dorsal ramus of the lacrimal is identical to that of T. rex and does not, as reported by Molnar (1980) resemble that of Albertosaurus ( Fig. 10 View Figure 10 ). We did not find the supraoccipital, but the median portion of the nuchal crest of the parietal is preserved ( Fig. 15J- M View Figure ). We regard the asymmetry in thickness between each ala of the nuchal crest as individual variation, in contrast to Molnar (1980) who regarded this as a shared similarity with Albertosaurus libratus . Likewise, we regard the absence of a medial shelf along the ventral margin of the surangular process of the dentary as individual variation.
We found that the olecranon process of the ulna is completely broken off and missing ( Fig. 14E, F View Figure 14 ), accounting for there being ‘no sign of a pronounced olecranal process as in Albertosaurus and Daspletosaurus ’ ( Molnar, 1980: 105).
Molnar listed similarities between LACM 23845 View Materials and Albertosaurus . Molnar stated that in LACM 23845 View Materials , ‘ A. lancensis ’, and A. libratus , the sagittal crest reaches the dorsal margin of the nuchal crest, but in Tyrannosaurus the crest does not reach the dorsal margin of the nuchal crest. However, among most tyrannosaurids ( Table 3 View Table 3 ), the extent of the sagittal crest is variable in most taxa and in LACM 23845 View Materials the sagittal crest actually stops short of the dorsal margin of the crest ( Fig. 15M View Figure ). Therefore, this character is individually variable and does not have diagnostic value except for A. libratus , in which the crest reaches the dorsal margin of the nuchal crest ( Table 3 View Table 3 ).
The form of the distal end of the fibula does not differ in any significant way from that of other tyrannosaurids. Molnar’s comparison of the metatarsals with Albertosaurus – in contrast to the stout bones in Tyrannosaurus – may simply reflect the smaller size of LACM 23845 View Materials . The supposed manual ungual is actually from digit I of the pes, based on the robust nature of the ungual and the low flexor tubercle. The angular form of the joint surface of the ungual is, in part, an artefact of breakage (cf. Molnar, 1980).
PREVIOUS TAXONOMIC WORK ON LACM 23845 View Materials
Paul (1988b: 324). referred LACM 23845 View Materials to the new species Albertosaurus megagracilis on the basis of its ‘extremely atrophied forelimbs, down-bent nasals, very long snout... long hind limbs’ and its ‘overall large size and gracile build’. With regard to the long snout, this is based on Molnar’s inference of a longer snout than in CMNH 7541 based on the greater (25%) length of the nasals in contrast to the slightly greater (10%) length of the frontals and parietals in LACM 23845 View Materials . Without an independent estimate of body size, it is difficult to determine what allometric trends these differences describe, and Paul’s (1988b) use of them as evidence of a taxonomic difference is undermined. Because LACM 23845 View Materials was shown above to be a subadult T. rex , A. megagracilis is a junior subjective synonym of T. rex .
Olshevsky & Ford (1995) state that LACM 23845 View Materials is not complete enough to be diagnosable but proceed to refer the specimen to a new taxon that they diagnose. Their diagnosis repeats characters from Molnar (1980) and they add the wide caudal region of the frontals, which indicates the orbits faced rostrally, but not as much in Tyrannosaurus . However, the caudal region of the frontal lies within the dorsotemporal fossa and is independent of orbit orientation ( Figs 8 View Figure 8 , 15C, D View Figure ). Olshevsky and Ford claim that there is a wide gap between the lacrimal and postorbital joint surfaces. However, the joint surface of the postorbital is missing on both sides, while a wide gap does not occur in subadult and adult tyrannosaurids (e.g. AMNH 5336, AMNH 5664, CMN 2120, ROM 1247). Unlike Olshevsky & Ford (1995), we suggest that LACM 23845 View Materials is diagnosable and can be referred to T. rex . Thus, D. megagracilis is a junior subjective synonym of T. rex .
DISCUSSION
We emphasize that our referral of LACM 28471 View Materials and LACM 23845 View Materials to T. rex is based on the presence of diagnostic characters of that species in both specimens. The stratigraphic position and geographical location of the specimens are consistent with the identifications; it is also most parsimonious to assume that small tyrannosaurids from late Maastrichtian beds are likely to be juvenile T. rex . The juvenile characters we document in both specimens are taxonomically neutral and they must be removed from the diagnoses of Aublysodon , Stygivenator and Dinotyrannus . If it were not for the presence of diagnostic characters in LACM 28471 View Materials and LACM 23845 View Materials , these specimens would be considered indeterminate small tyrannosaurids because several of the juvenile characters also typify most adult small theropods (e.g. dorsoventrally shallow dentaries; Table 4 View Table 4 ) and so do not support our argument because they are not diagnostic in and of themselves.
CRANIOFACIAL ONTOGENY IN
The hypothesis that LACM 28471 View Materials and LACM 23845 View Materials are juvenile and subadult specimens, respectively, of T. rex adds to our knowledge of growth in this taxon (cf. Carr, 1999).
Figure 17. Continued
QUANTITATIVE ONTOGENETIC ANALYSIS A quantitative analysis of 84 morphological characters (see Appendix 1) among five individuals of T. rex and a hypothetical embryo was performed using PAUP v. 3.1 ( Swofford, 1993). The data matrix ( Appendix 2) was compiled using MacClade v. 3.0 ( Maddison & Maddison, 1992) and then run in PAUP under ACCTRAN and DELTRAN optimizations. One hundred replicates were run under a heuristic search using a random addition sequence, tree-bisection-reconnection branch swapping, with the MULPARS option in effect. Finally, the trees were unrooted and multistate individuals were treated as polymorphisms. The characters were polarized by constructing a hypothetical embryo in which all states were coded as nascent and coded in a fashion analogous to the primitive state of a phylogenetic character (i.e. as ‘0’). The embryo was specified as the outgroup. The oldest specimen was specified using an artificial adult of the most mature characters. The specimen placed in a ‘sister species’ position with the artificial adult was considered to be the oldest adult.
Analysis of the ontogenetic characters resulted in a single branching diagram that arrayed the five specimens along an ontogenetic gradient or growth series ( Fig. 19 View Figure 19 ). The tree is 89 steps long and, excluding uninformative characters, has a consistency index (CI) of 0.97, a homoplasy index (HI) of 0.03, and retention index (RI) of 0.97. Five growth stages can be described in terms of unambiguous character transformations (see Appendix 3).
In the first stage (small juvenile, e.g. LACM 238471 View Materials ), the rostroventral margin of the antorbital fossa grades into the lateral surface of the maxilla ( Figs 1D View Figure 1 , 19 View Figure 19 ). In small juvenile specimens of Albertosaurus libratus , this margin is distinct and is presumably the juvenile condition. Also, in small juveniles of T. rex the first maxillary tooth is nondenticulate and, in ventral view, the joint surface for the internasal septum is wide, indicating that the septum was disproportionately wide in juveniles ( Fig. 1C View Figure 1 ). In contrast to larger specimens (e.g. CMNH 7541 ), the sagittal ridge that overlies the closed internasal suture is relatively wide, occupying almost one-third the width of the paired bones ( Fig. 1A, B View Figure 1 ). In dorsal and ventral views, the interparietal suture is closed, although it is traceable rostrally in ventral view ( Fig. 2L, M View Figure 2 ). The nuchal crest of small juveniles is only moderately indented on the midline in caudal view ( Fig. 2P View Figure 2 ). In larger specimens ( CMNH 7541 ), this indentation is deep and the sagittal crest projects above the dorsal margin as a triangular projection in caudal view ( Carr, 1999). LACM 28471 View Materials appears to indicate the polarity of the frontoparietal suture in derived tyrannosaurid evolution: the suture is V-shaped in dorsal view, indicating that the transverse condition of Daspletosaurus and Tyrannosaurus may be derived, while the wedgelike suture of Albertosaurus might be primitive.
In the second growth stage (large juvenile, e.g. CMNH 7541 ), the sagittal crest is present and deep on the frontal, the frontoparietal suture is transverse, and the dentary is deep ( Fig. 19 View Figure 19 ). The greatest morphological development occurs between stages two and three (subadult, e.g. LACM 23845 View Materials ). This is due in part to the lack of specimens to fill the gap between the juvenile and subadult specimens included in the analysis ( Fig. 19 View Figure 19 ). Cranial ornamentation is elaborated in subadults, such as the appearance of gnarled nasals ( Fig. 19 View Figure 19 ). Changes necessary for a mechanically optimal large skull are implied by evidence of intracranial buttressing, such as columnar and thick nasals and a deep frontal and parietal ( Figs 9 View Figure 9 , 15A, B, F, G View Figure , 19 View Figure 19 ). Inflation of bones by the antorbital air sac is reflected by the constricted frontal process of the nasals between the lacrimals, and the short and wide frontal ( Figs 8 View Figure 8 , 9B View Figure 9 , 15C, D View Figure ). Increase in the mass of the temporal musculature is indicated by the deep dorsotemporal fossa on the frontal ( Figs 8 View Figure 8 , 15A, C, D View Figure ), low and wide sagittal crest of the parietal ( Fig. 15G, H View Figure ), lateroventral orientation of the surangular shelf ( Fig. 17A View Figure 17 ), deep surangular and prearticular ( Fig. 17A, B, D, F View Figure 17 ), rostral position of the dorsal margin of the prearticular ( Fig. 17D, F View Figure 17 ), and caniniform teeth. Also, the thick nuchal crest reflects hypertrophied cervical musculature ( Fig. 15J- M View Figure ). Other changes are less easily accounted for, such as the enlarged marginal alveolar foramina of the maxilla ( Fig. 11A View Figure 11 ), elongate and laterally concave premaxillary process of the nasal ( Fig. 9B, D View Figure 9 ), and deep and rugose scar lateroventral to the glenoid fossa of the surangular ( Fig. 17A View Figure 17 ).
In the fourth stage (young adult, e.g. AMNH 5027 View Materials ), the ridge that encircles the rostroventral margin of the antorbital fossa is obliterated by the thickened nature of the maxilla, the rostral ramus of the lacrimal is grossly inflated such that the ramus is deeper than the pneumatic recess, and the surangular shelf is orientated horizontally ( Fig. 19 View Figure 19 ). Although the analysis including the artificial adult was equivocal between the two adult specimens, the fifth stage (old adult) is logically represented by LACM 23844 View Materials because it exhibits additional pneumatic destruction of bone in the antorbital fossa, principally in the interfenestral strut ( Fig. 19 View Figure 19 ).
DENTICLE SIZE DIFFERENCE INDEX (DSDI)
Rauhut & Werner (1995) proposed DSDI as a sizeindependent means of comparing denticle densities (number of denticles per given unit length) among theropods. A purview of the DSDIs obtained for various growth stages of T. rex ( Table 5 View Table 5 ) indicates that DSDIs decrease in progressively larger specimens, that is, there are fewer mesial denticles per given unit length than distal denticles in large specimens and there are as many or more mesial than distal denticles in small specimens. Also, the DSDI among dentary teeth is higher than that in the maxilla (e.g. LACM 28471 View Materials ; Table 5 View Table 5 ), indicating that mesial denticles are smaller in the dentary (1.20) than in the maxillary (1.04) dentition.
DISCUSSION
PREMAXILLARY TOOTH VARIATION IN
TYRANNOSAURIDS
Lehman & Carpenter (1990) consider smooth carinae that converge at midheight of the crown and a bilobed median ridge on the lingual surface to diagnose the premaxillary teeth of Aublysodon . The authors consider denticulate and parallel carinae to be characteristic of other tyrannosaurids. It appears this observation is limited to Leidy’s (1860) figures of Deinodon where it is clear that the large denticulate crowns are missing the basal region where the carinae converge (1860, pl. 9, figs 35–40). Also, the premaxillary teeth of a new basal tyrannosauroid (NMMNH P-25049) and cf. Albertosaurus sp. (ROM 1422) display basally converging, denticulate carinae. Although they do not list it as a diagnostic character, Lehman & Carpenter (1990) also note a bilobed median ridge on the lingual surface of the tooth; this ridge is typical among tyrannosaurids, including cf. Albertosaurus sp. (ROM 1422) and a new genus of tyrannosauroid (NMMNH P-25049).
CONCLUSIONS
Based on the evidence presented herein, it is most parsimonious to identify LACM 28471 View Materials as a small juvenile T. rex and LACM 23845 View Materials as a subadult T. rex instead as different genera. Aublysodon is an invalid taxon; the skeletal characters ascribed to it are either plesiomorphic among Tyrannosauridae or growth-related. Likewise, Stygivenator molnari and Dinotyrannus megagracilis are invalid. Although Carpenter has suggested the Tornillo Formation of Texas has a large theropod that is not T. rex , it is referable to T. rex ( Carr & Williamson, 2000).
LATE MAASTRICHTIAN TYRANNOSAURID DIVERSITY
Currently, there is evidence for only one tyrannosaurid species, T. rex , in late Maastrichtian strata of western North America. This contrasts with the diversity in the late Campanian, where the occurrence of two genera in the same unit is not uncommon (e.g. Albertosaurus and Daspletosaurus in the Dinosaur Park Formation; Russell, 1970). Also during the late Campanian, different tyrannosauroid species were present in different regions of the west at approximately the same time.
DSDI
The variation in DSDI in T. rex indicates that individual and ontogenetic variation in this index must be defined before it is used to supplement taxonomic referrals (e.g. Rauhut & Werner, 1995). For example, the DSDI in the juvenile T. rex LACM 28471 View Materials matches that of a tooth (1.33) referred to Dromaeosauridae ( Rauhut & Werner, 1995) . The total range of DSDIs for the growth series is 0.62–1.33, which virtually encompasses that obtained by Rauhut & Werner for Theropoda, 0.60–2.33 (1995); when theropods with apomorphically fine mesial denticles are excluded (Velociraptorinae, Richardoestesia ) the range is 0.60– 1.20, which is encompassed by the variation in T. rex .
SIGNIFICANCE
LACM is currently the only museum that holds a growth series of T. rex . LACM 23845 View Materials helps to fill in the gaps in the growth series between the meter-long skulls of adults and the 60-cm skull of CMNH 7541 . LACM 28471 View Materials extends our knowledge of the morphology of small T. rex juveniles.
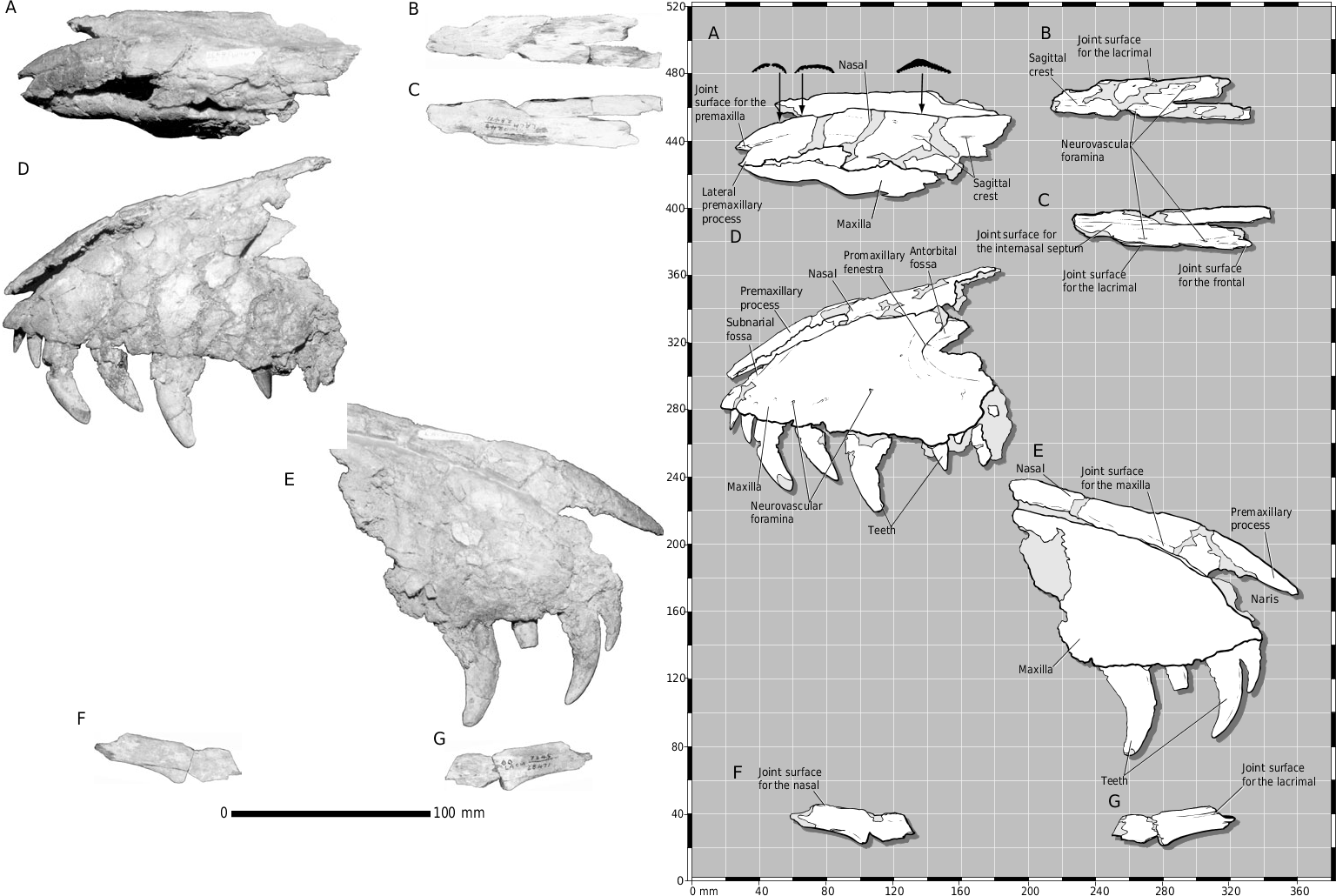
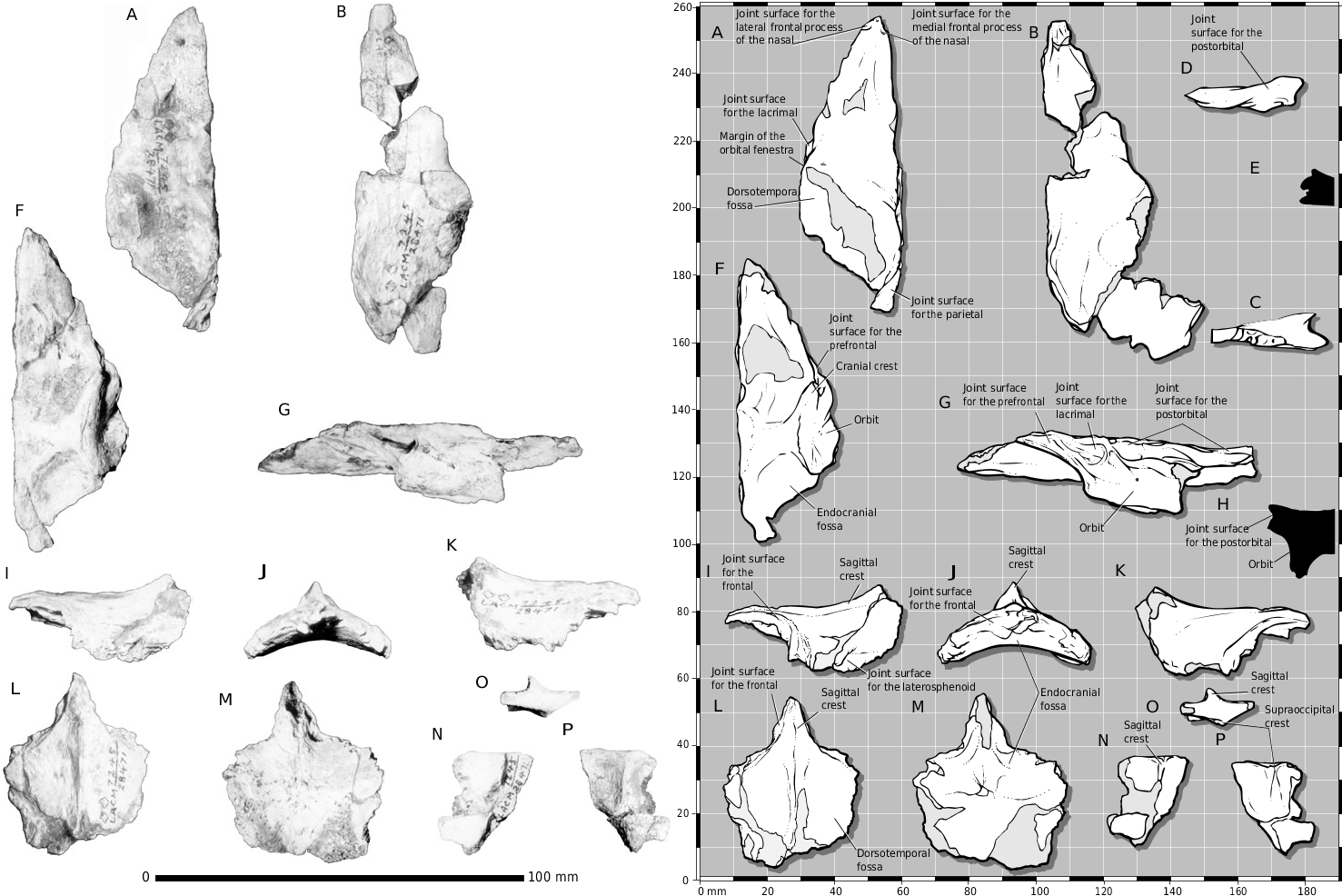
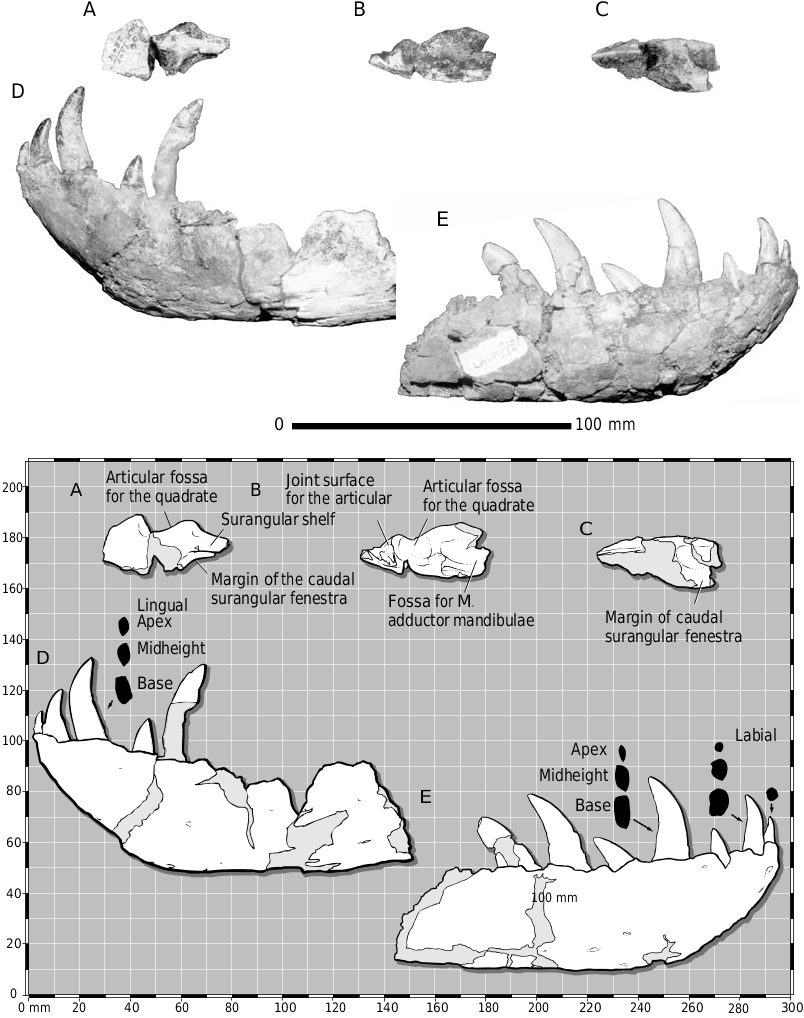



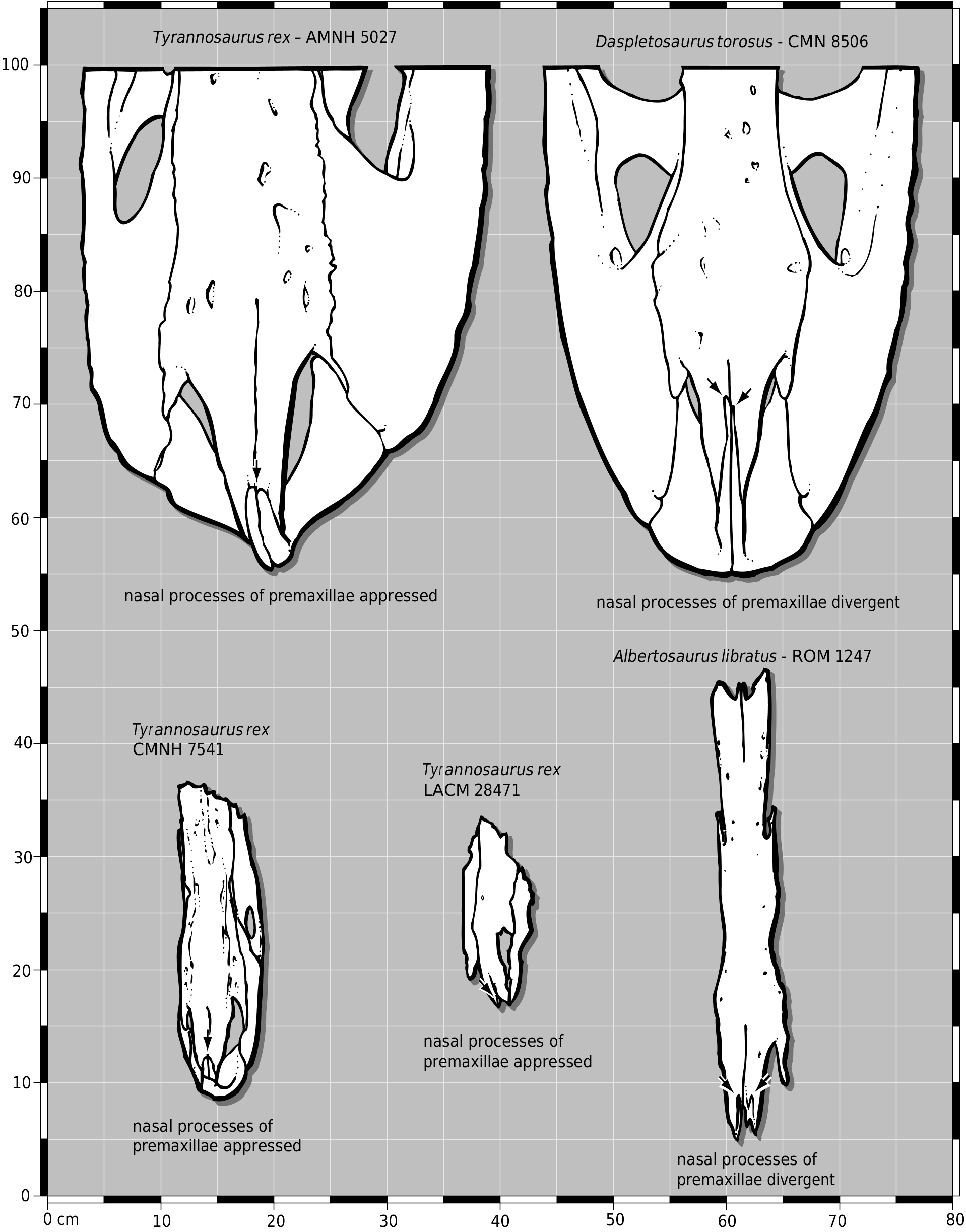


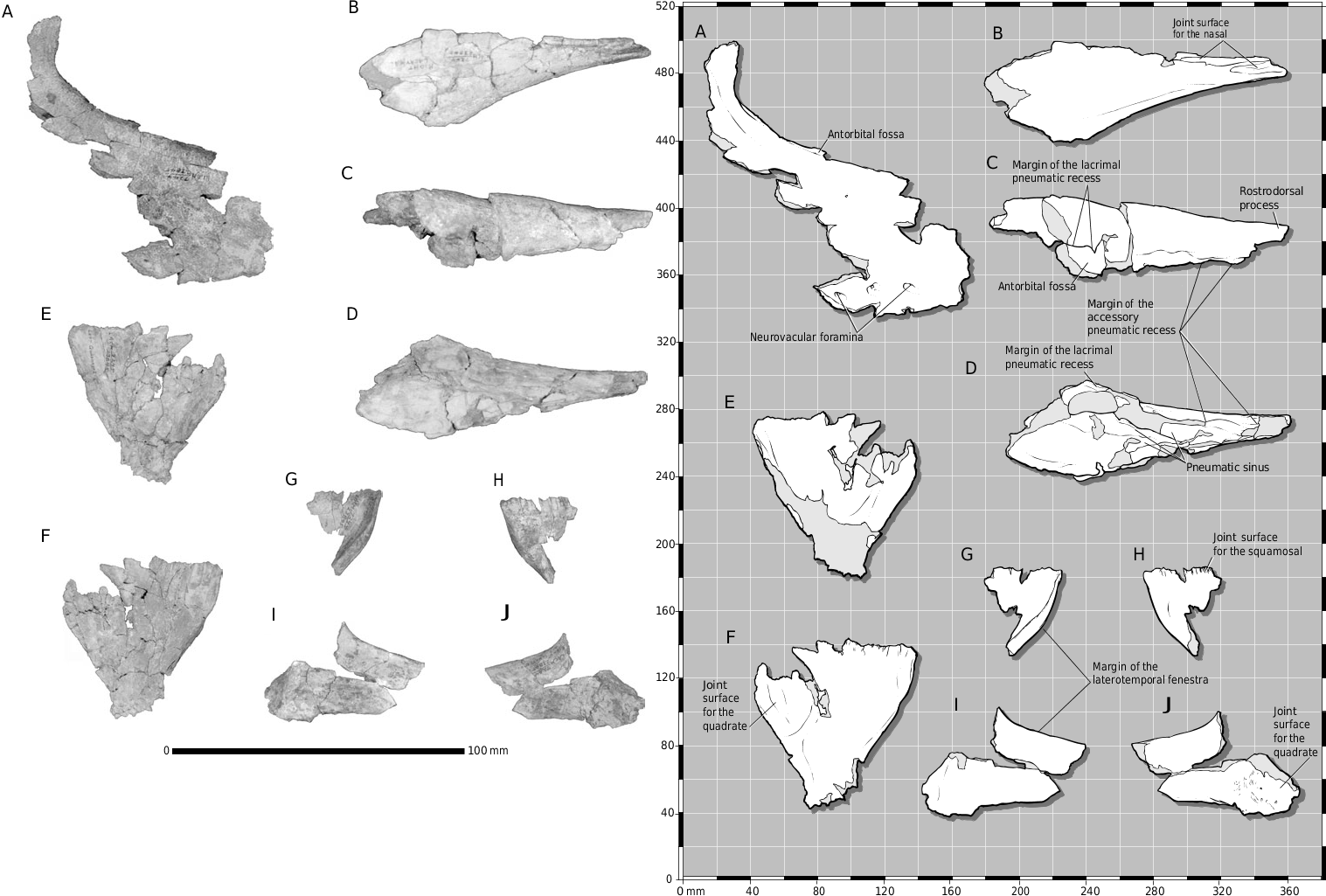
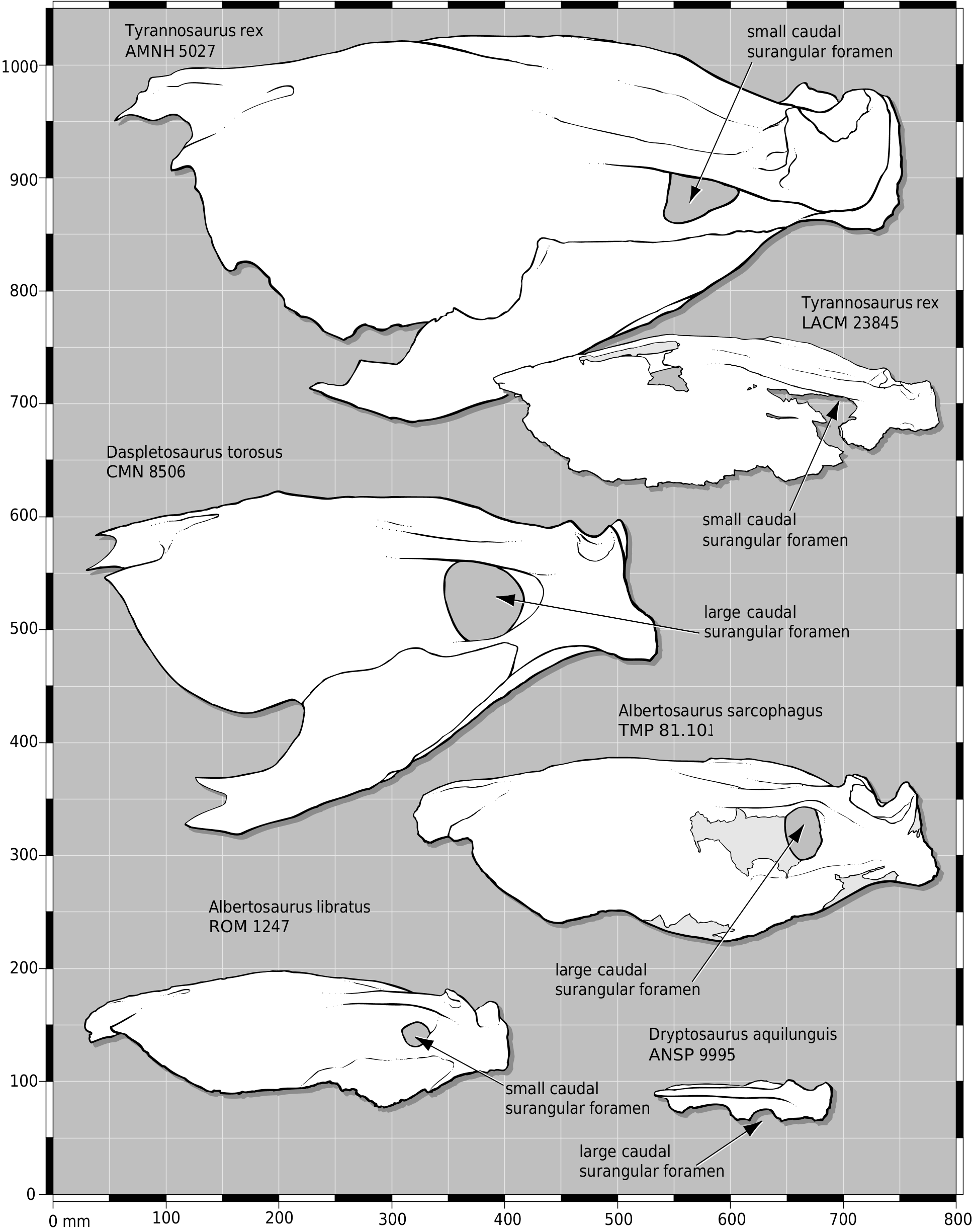

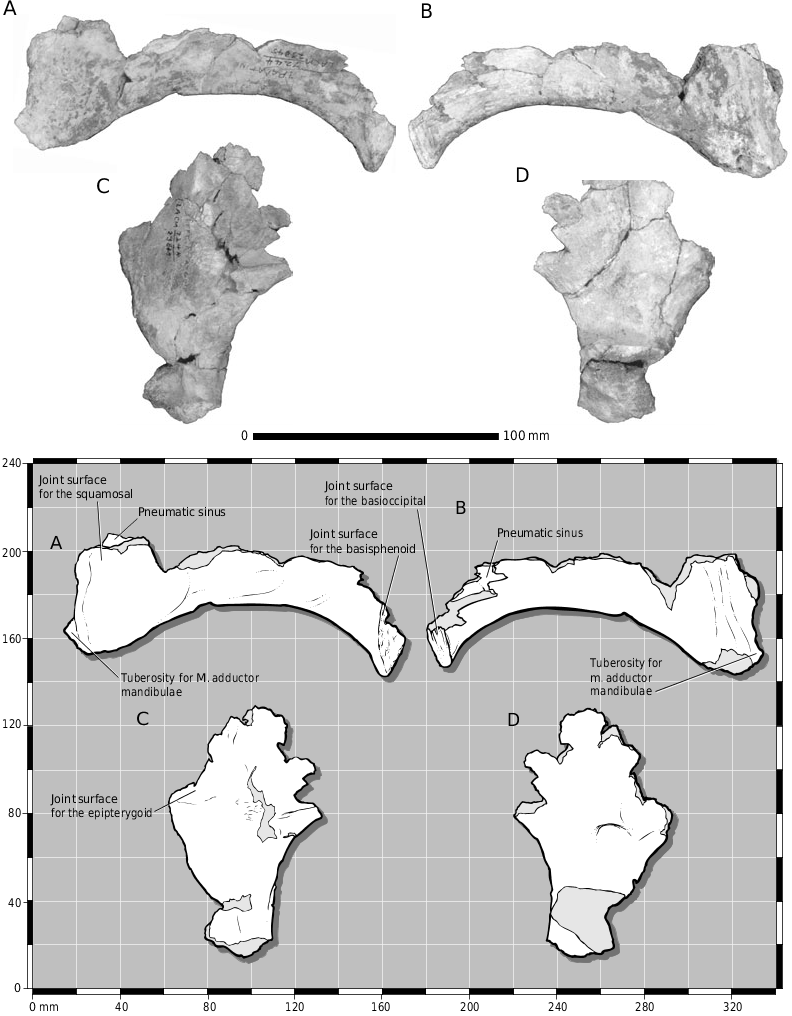


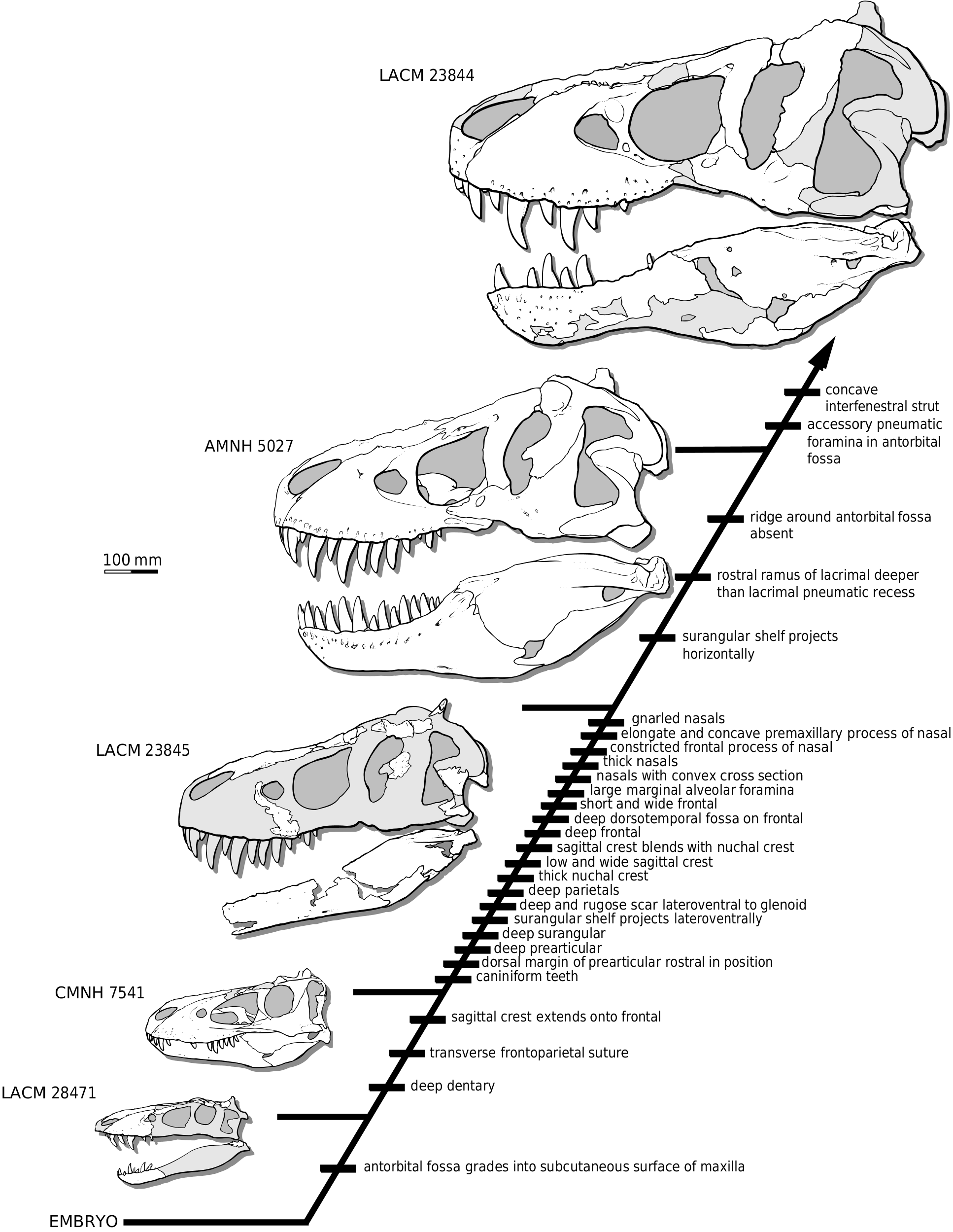
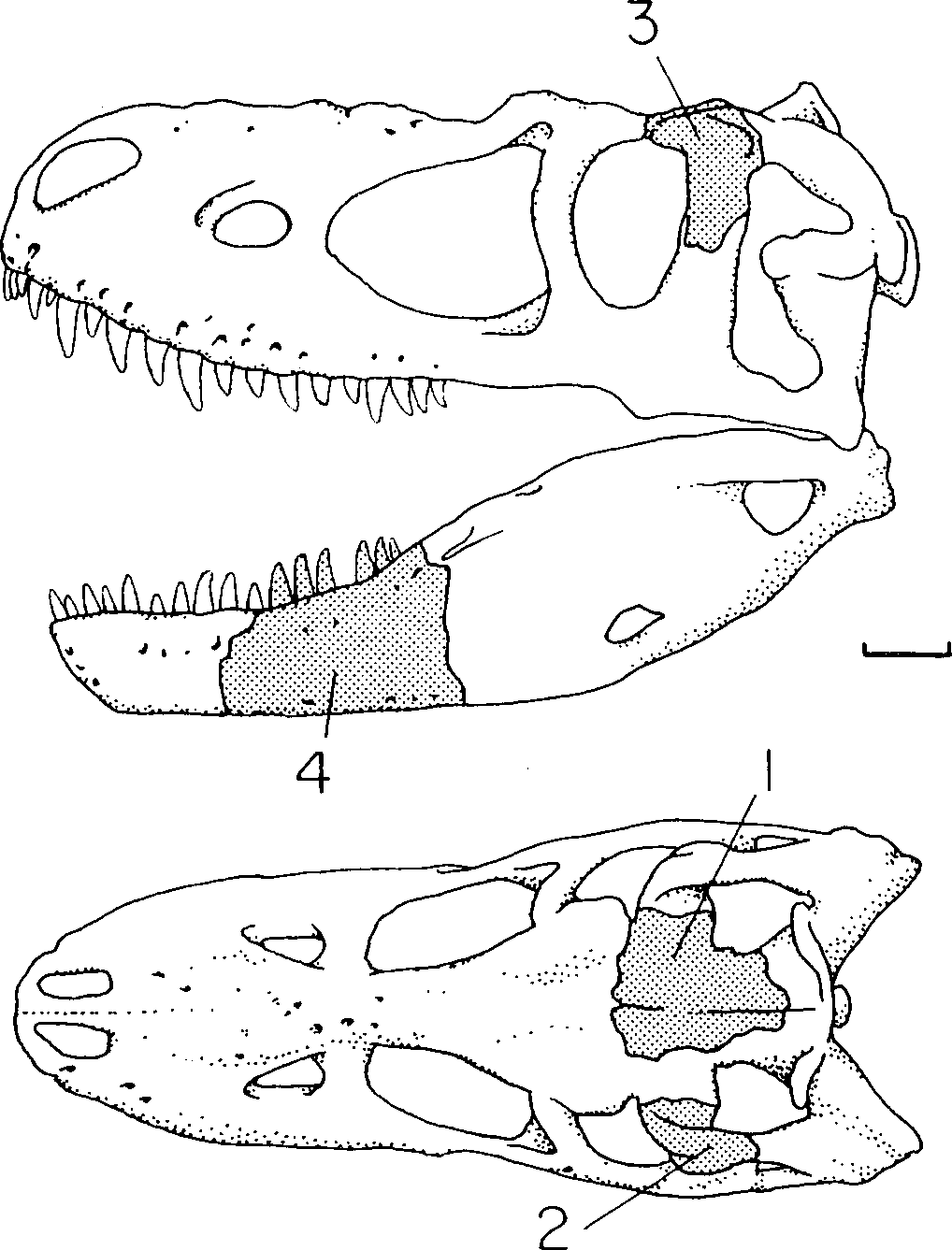

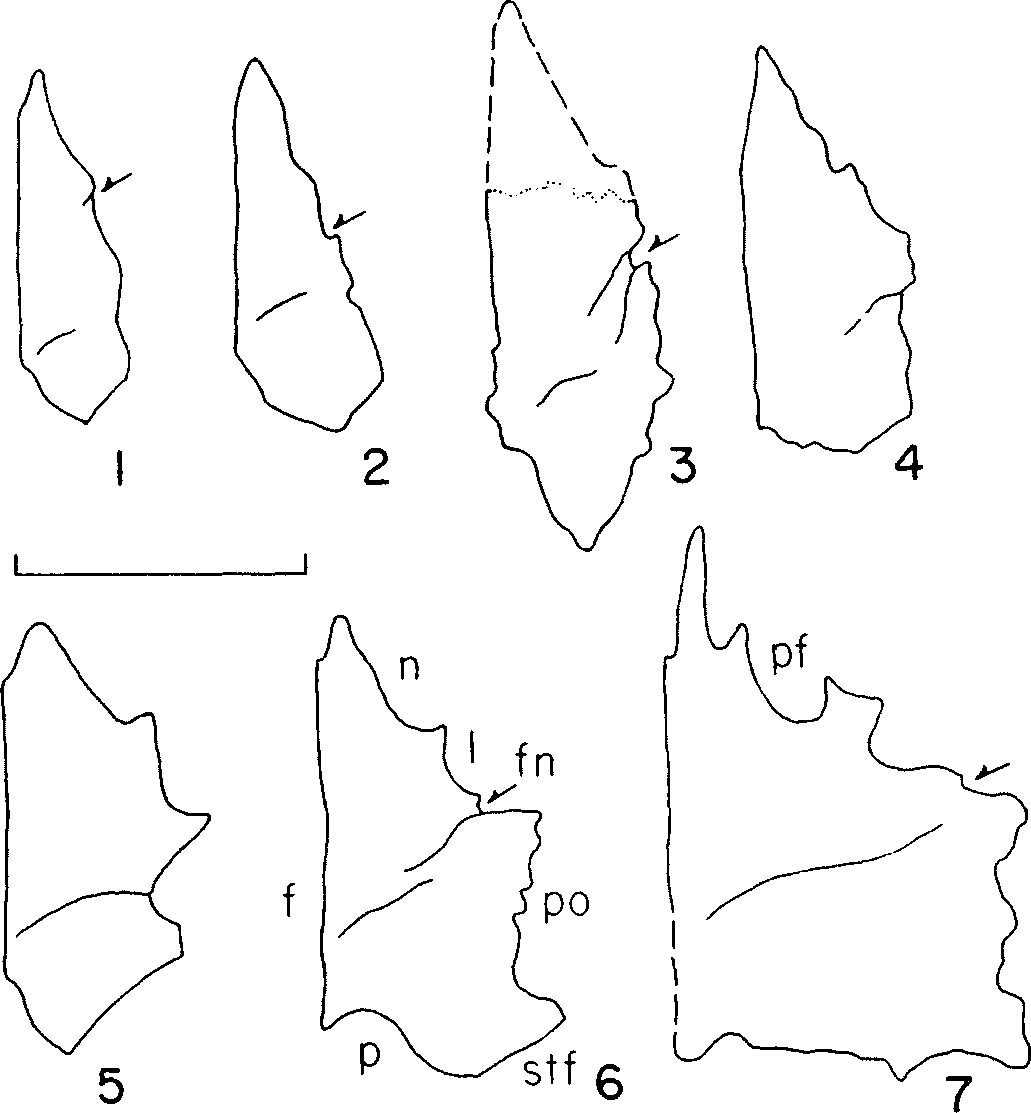
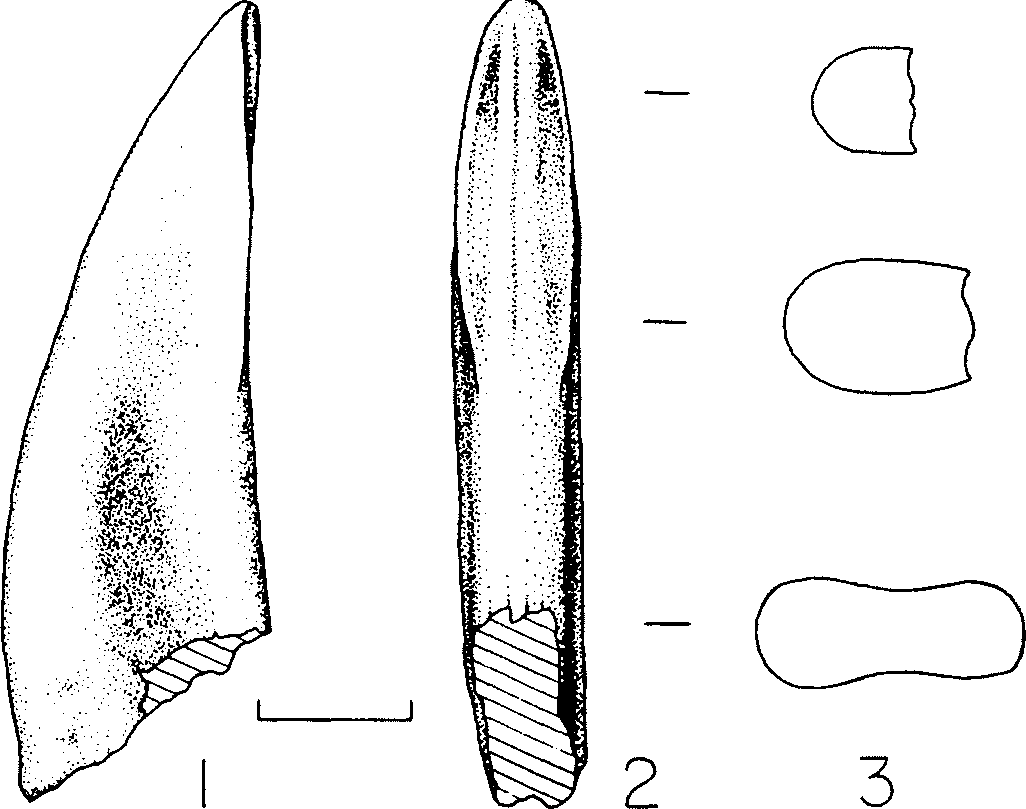


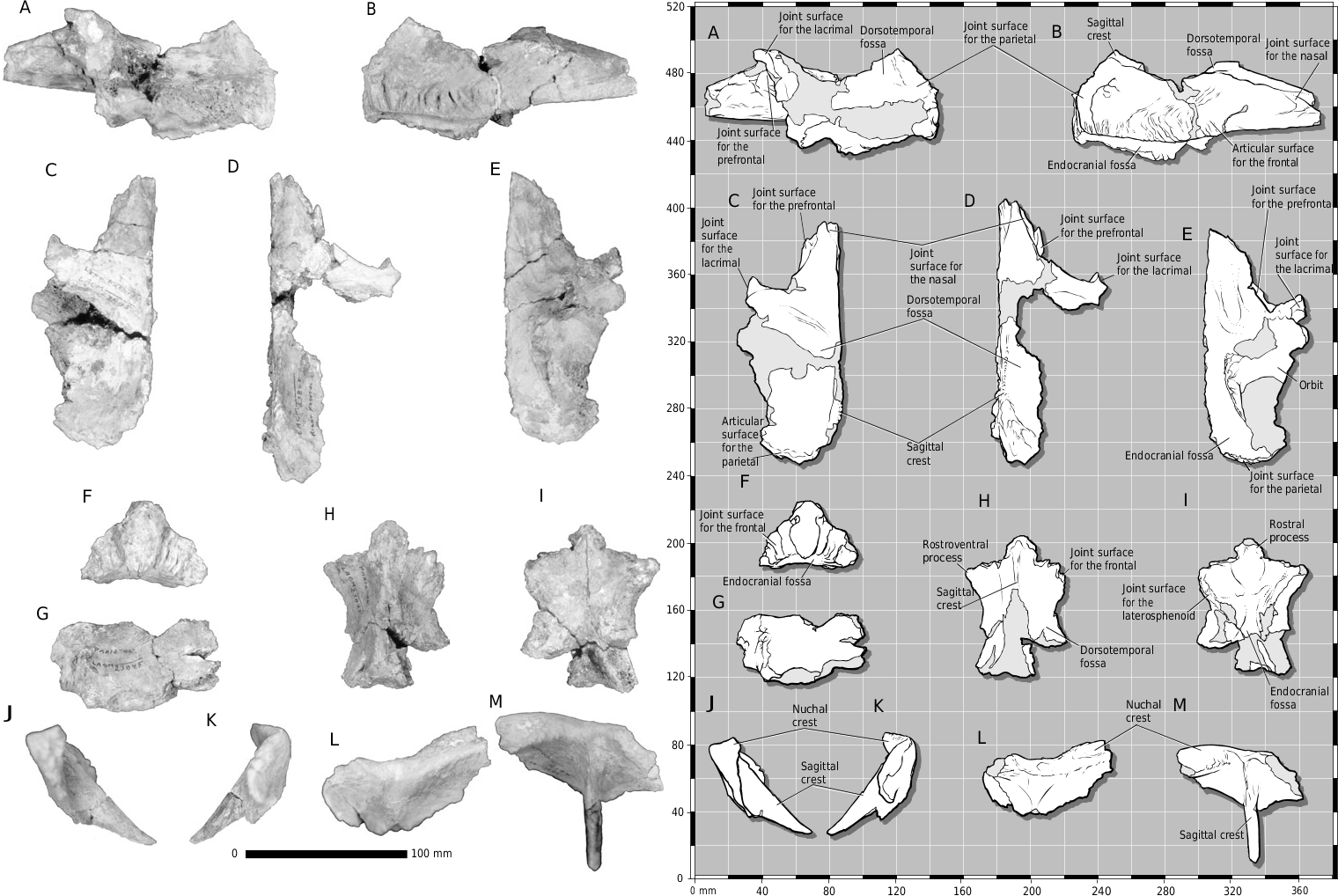
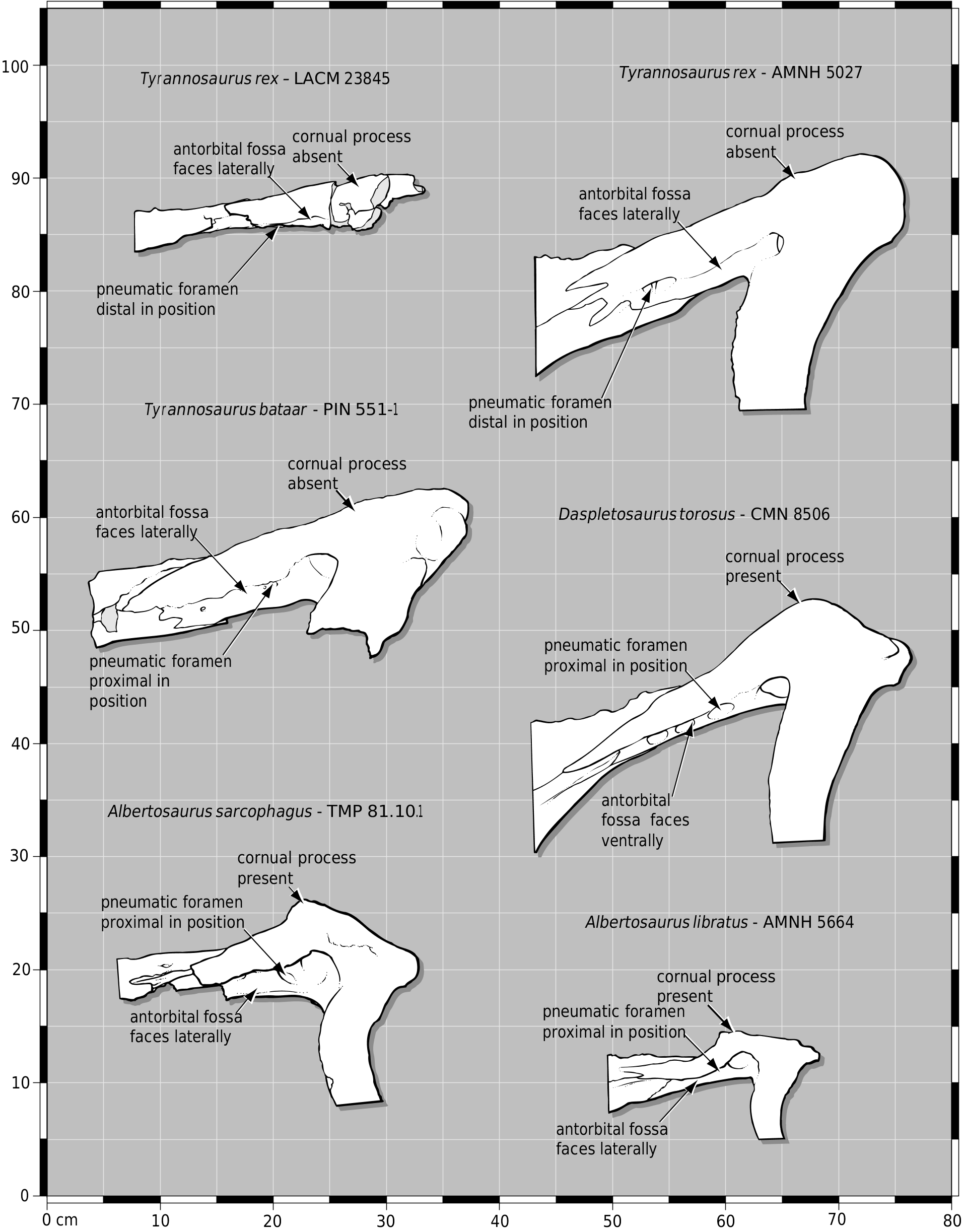

Table 1. Differences in opinion of the osteology of LACM 28471 between previous authors and the interpretations presented in this paper
| Molnar (1978) | Molnar & Carpenter (1989) | Olshevsky & Ford (1995) | This paper |
|---|---|---|---|
| – | Emarginated dentary | – | Damaged transition region |
| Dorsotemporal fossa absent | – | – | Dorsotemporal fossa present |
| Tract of subnarial foramen may be preserved | – | – | Fossa of subnarial foramen is preserved |
| Nasals shallow | – | – | Nasal deep, but crushed |
| Frontoparietal suture serrate | – | – | Frontoparietal suture serrate laterally |
| Joint surface of laterosphenoid on the frontal | – | – | Joint surface of the postorbital on the frontal |
| Joint surface of the prefrontal on the frontal | – | – | Joint surface of the lacrimal on the frontal |
| Absence of ossification of the crista cranii of the frontals with other bones | – | – | Disarticulation and loss of the prefrontal from the frontal |
| Joint surface of the prootic on the parietal | – | – | Joint surface of the laterosphenoid on the parietal |
| Lateral buttress of the surangular | – | – | Surangular shelf |
| Meckelian groove absent | – | – | Meckelian groove present |
| Interdental plates absent | – | – | Interdental plates present |
| Premaxillary tooth present | Premaxillary tooth present | Premaxillary tooth present | First maxillary tooth |
| D-shaped first dentary tooth | – | – | Conical first dentary tooth |
| – | Caudolateral margin of frontal absent | – | Caudolateral margin of frontal present |
| Joint surface of the postorbital on the frontal is absent | – | Joint surface of the postorbital on the frontal is present | |
| – | Joint surface of the prefrontal on the frontal | – | Joint surface of the lacrimal on the frontal |
| – | Region of dorsotemporal fossa flat | – | Dorsotemporal fossa concave |
| – | V-shaped rostral margin of antorbital fossa | – | Rostral margin of antorbital fossa extends caudodorsally |
Table 2. Differences in opinion regarding the osteology of LACM 23845 between Molnar (1980) and this paper
| Molnar (1980) | This paper | |
|---|---|---|
| Nasals | flexedrostroventrally | damaged |
| Frontal process of nasal | not strongly constricted | incomplete but constricted |
| Dorsal ramus of lacrimal | shallow | only upper half present |
| Joint surface of the postorbital on the frontals | present | absent |
| Joint surface of the prefrontal on the frontal | smoothly concave | rugose |
| Frontoparietal suture | V-shaped | transverse |
| Supraoccipital | present | notpreserved |
| Olecranon process of ulna | apomorphically absent | broken off and missing |
| Ungual | manual ungual present | Pedal D I ungual |
| Joint facet of ungual | angular | damaged |
Table 3. Variation of the position and width of the end of the sagittal crest on the nuchal crest among tyrannosaurids. The number of specimens exhibiting a given state is in parentheses
| Specimen/ Taxon | Position | Width | Variation |
|---|---|---|---|
| Albertosaurus libratus | |||
| AMNH 5336 | reaches dorsal margin | narrow | reaches dorsal margin; narrow (3) or |
| ROM 1247 | reaches dorsal margin | wide | wide (1) |
| TMP 86.144.1 | – | narrow | |
| TMP 91.36.500 | reaches dorsal margin | narrow | |
| A. sarcophagus | |||
| AMNH 5222 | stops short | narrow | stops short (2) or reaches (1) |
| TMP 81.10.1 | stops short | narrow | dorsal margin; narrow (2) or wide (2) |
| TMP 85.98.1 | reaches dorsal margin | wide | |
| TMP 86.64.1 | – | wide | |
| Daspletosaurus | |||
| CMN 8506 | reaches dorsal margin | narrow | stops short (2) or reaches (4) dorsal |
| CMN 11594 | stops short | wide | margin; narrow (4) or wide (1) |
| FMNH PR308 | reaches dorsal margin | narrow | |
| TMP 85.62.1 | reaches dorsal margin | narrow | |
| TMP 94.143.1 | – | narrow | |
| New genus | |||
| NMMNH P-25049 | reaches dorsal margin | wide | |
| NMMNH P-27469 | stops short | narrow | stops short of dorsal margin or reaches; narrow or wide |
| Tyrannosaurus rex | |||
| AMNH 5027 View Materials | stops short | wide | stops short (4) or reaches (5) dorsal |
| AMNH 5029 View Materials | reaches dorsal margin | narrow | margin; narrow (8) or wide (1) |
| AMNH 5117 View Materials | reaches dorsal margin | narrow | |
| BHI 3033 | reaches dorsal margin | narrow | |
| CMNH 7541 | reaches dorsal margin | narrow | |
| FMNH PR2081 | stops short | narrow | |
| LACM 23845 View Materials | stops short | narrow | |
| LACM 28471 View Materials | reaches dorsal margin | narrow | |
| TMP 81.6.1 | stops short | narrow |
Table 4. Comparison of the juvenile characters of LACM 28471 and LACM 23845 with adults of selected clades of small late Cretaceous theropods from various sources: Troodontidae (Barsbold, 1974; Currie, 1985, 1987, 1995; Currie & Dong, 2001b; Xu et al., 2002); Dromaeosauridae (Ostrom, 1969; Sues, 1978; Currie, 1987; Xu & Wu, 2001; AMNH 6515 (cast); PIN 3143/8 (cast); MOR 747); Ornithomimidae (Currie, 1987; ROM 840; TMP 90.26.1; Parks, 1928; Osmolska et al., 1972; Barsbold, 1983)
| LACM 28471 View Materials | Troodontidae | Dromaeosauridae | Ornithomimidae | |
|---|---|---|---|---|
| Teeth | nondenticulate | nondenticulate and denticulate | nondenticulate and denticulate | nondenticulate |
| narrow lateral | narrow lateral | narrow lateral | narrow lateral | |
| Skull bones | lightly built | lightly built skull | lightly built | lightly built |
| Nasomaxillary suture | smooth | smooth | smooth | smooth |
| Lateral margins of frontal process of nasal | parallel | parallel | parallel | parallel |
| Maxilla | shallow and narrow | shallow and narrow | shallow and narrow | shallow and narrow |
| Maxillary fenestra | caudal to rostral margin of antorbital fossa | caudal | caudal | caudal and at the rostral margin |
| Rostral margin of dorsotemporal fossa | indistinct | distinct | distinct | indistinct |
| Joint surface of postorbital of frontal | series of grooves and ridges | pair of facets | pedunculate | - |
| Joint surface of the lacrimal on the frontal | long and narrow | long and narrow | short and narrow and short and wide | – |
| Joint surface of the prefrontal on the frontal | long and narrow | prefrontal absent? | long and narrow | long and narrow |
| Frontal processes of the nasal | exposed dorsally | exposed dorsally | exposed dorsally | exposed dorsally |
| Scar lateroventral to glenoid fossa of the | ||||
| surangular | shallow groove | – | absent | – |
| Dentary | dorsoventrally shallow | dorsoventrally shallow | dorsoventrally shallow | dorsoventrally shallow |
| Dorsal and rostroventral margins of the dentary | meet at a low angle | meet at high and low angles | meet at high and low angles | meet at high and low angles |
| LACM 23845 View Materials | Troodontidae | Dromaeosauridae | Ornithomimidae | |
| Nasals | rugose | smooth and rugose | smooth | rugose |
| Frontonasal suture | W-shaped | V-shaped | W-shaped and V-shaped | V-shaped |
| Dorsal ramus of lacrimal | shallow | shallow | shallow | shallow |
| Nuchal crest | smooth | smooth | smooth | smooth |
| Tubercle for M. depressor mandibulae | ||||
| of the otoccipital | low | absent | absent | – |
Table 5. Comparison of DSDIs (Denticle Size Difference Indices) in a growth series of Tyrannosaurus rex showing sizeindependent variation. Denticles were counted per 5 mm at the base, midheight and apex of the carinae. Abbreviations: a, apical; b, basal; D, dentary; L, left; M, maxilla; m, midheight; R, dentary; U, unknown source bone; numerals indicate alveolus position
| Specimen | DSDIs | DSDI range and average |
|---|---|---|
| LACM 238471 View Materials | LM4m: 1.00m, 1.08a; LM6: 1.00a; RM4: 1.08b; RM5: 1.08m; LD3: 1.21m; RD3: 1.33a; RD4: 1.14a; RD5: 1.20a; RD7: 1.07m | Range: 1.00–1.33 Average: 1.12 |
| CMNH 7541 | LM3: 1.08m; RM7: 0.92a | Range: 0.91–1.08 Average: 1.00 |
| LACM 23845 View Materials | U: 0.92b, 1.11m, 0.90a; U: 0.93b, 0.83m; U: 1.00b, 0.94m | Range: 0.83–1.11 Average: 0.95 |
| NMMNH P-3698 | U: 0.75a, 0.80m, 0.86b; U: 0.86m; U: 0.86a, 0.88m, 0.62b; U: 1.00a | Range: 0.62–1.00 Average: 0.83 |
| Total range: 0.62–1.33 Total average: 0.95 |
| TMP |
Taiwan [Republic of China], Taipei, Taiwan Provincial Museum |
| LACM |
USA, California, Los Angeles, Los Angeles County Museum of Natural History |
| CMNH |
USA, Pennsylvania, Pittsburgh, Carnegie Museum of Natural History |
| AMNH |
USA, New York, New York, American Museum of Natural History |
| BHI |
BHI |
| FMNH |
USA, Illinois, Chicago, Field Museum of Natural History (also used by Finnish Museum of Natural History) |
| SDSM |
SDSM |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tyrannosaurus rex
| Thomas D. Carr & Thomas E. Williamson 2004 |
Stygivenator molnari (Paul, 1988b)
| Stygivenator molnari (Paul, 1988b) - Olshevsky & Ford, 1995 : fig. 36, p. 116 |
Dinotyrannus megagracilis Olshevsky & Ford, 1995
| Dinotyrannus megagracilis Olshevsky & Ford, 1995 : fig. 37, p. 117 |
Aublysodon molnaris (Leidy, 1868)
| Aublysodon molnaris (Leidy, 1868) - Paul, 1988b : p. 325 |
Aublysodon cf. A. mirandus (Leidy, 1868)
| Aublysodon cf. A. mirandus (Leidy, 1868) - Molnar & Carpenter, 1989 : figs 1-4, p. 447 |
Albertosaurus cf. A. lancensis
| Albertosaurus cf. A. lancensis Molnar, 1980 : figs 1-7, p. 102 |
Albertosaurus megagracilis
| Albertosaurus megagracilis Paul, 1988b : p. 333. |
Nanotyrannus lancensis
| Nanotyrannus lancensis Bakker, Williams, & Currie, 1988 : pls. 1-4, figs 1-7, 11, 12, p. 2 |