
Sabellomma collinae, Nogueira, João Miguel De Matos, Fitzhugh, Kirk & Rossi, Maíra Cappellani Silva, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.275967 |
|
DOI |
https://doi.org/10.5281/zenodo.6209683 |
|
persistent identifier |
https://treatment.plazi.org/id/03036267-800E-FFE2-FF6B-5451FB4EFE97 |
|
treatment provided by |
Plazi |
|
scientific name |
Sabellomma collinae |
| status |
sp. nov. |
Sabellomma collinae View in CoL spec. nov.
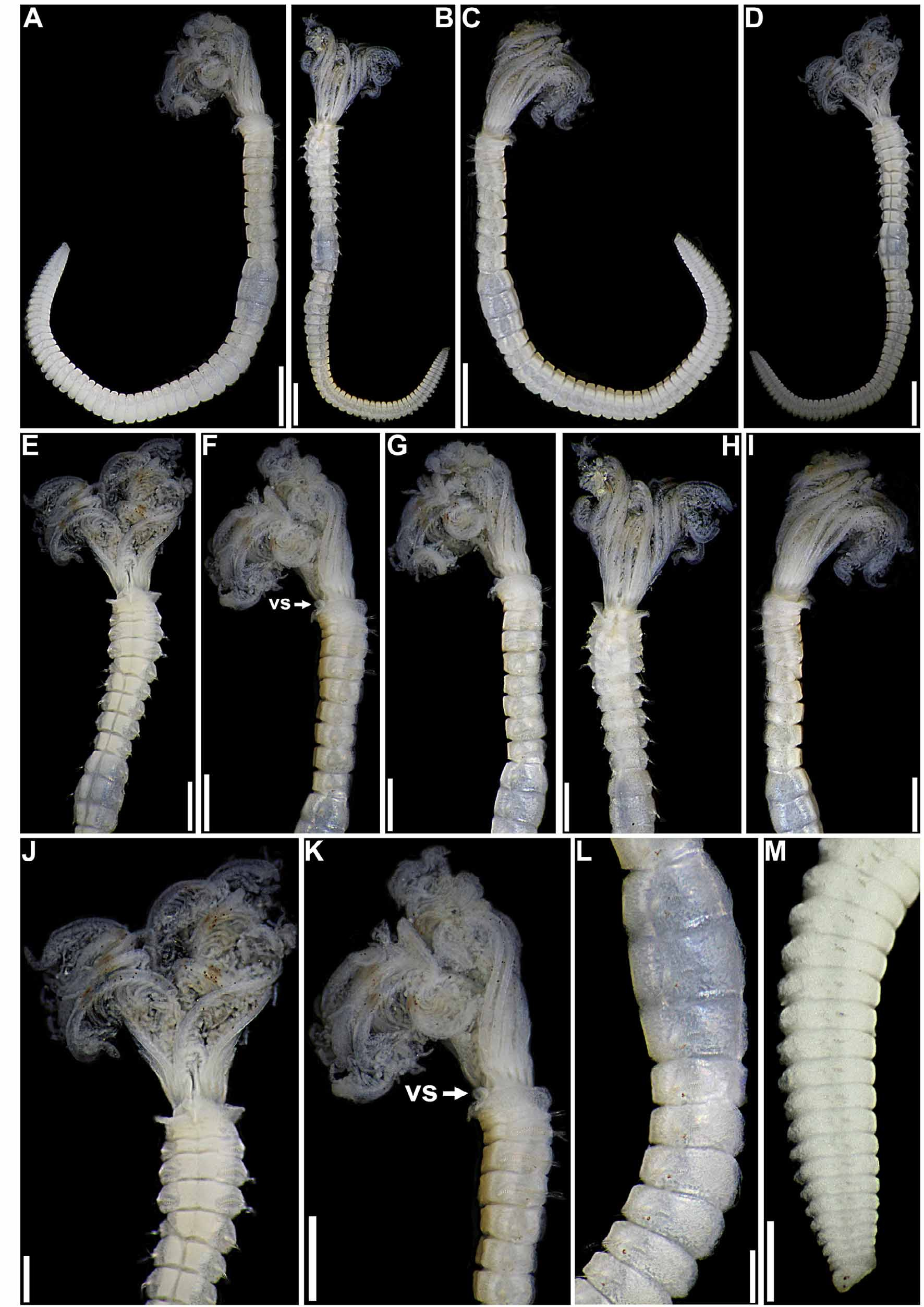
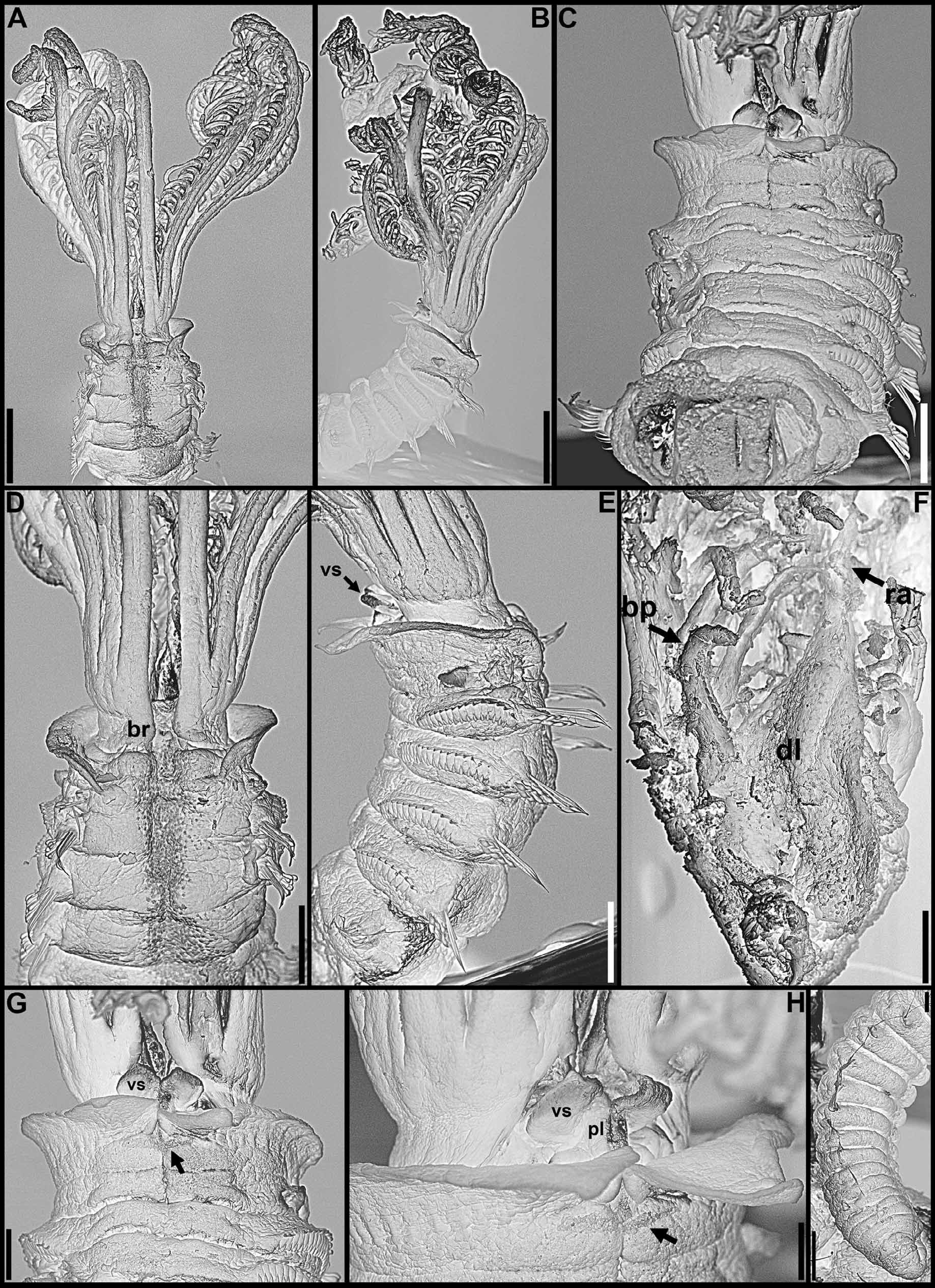
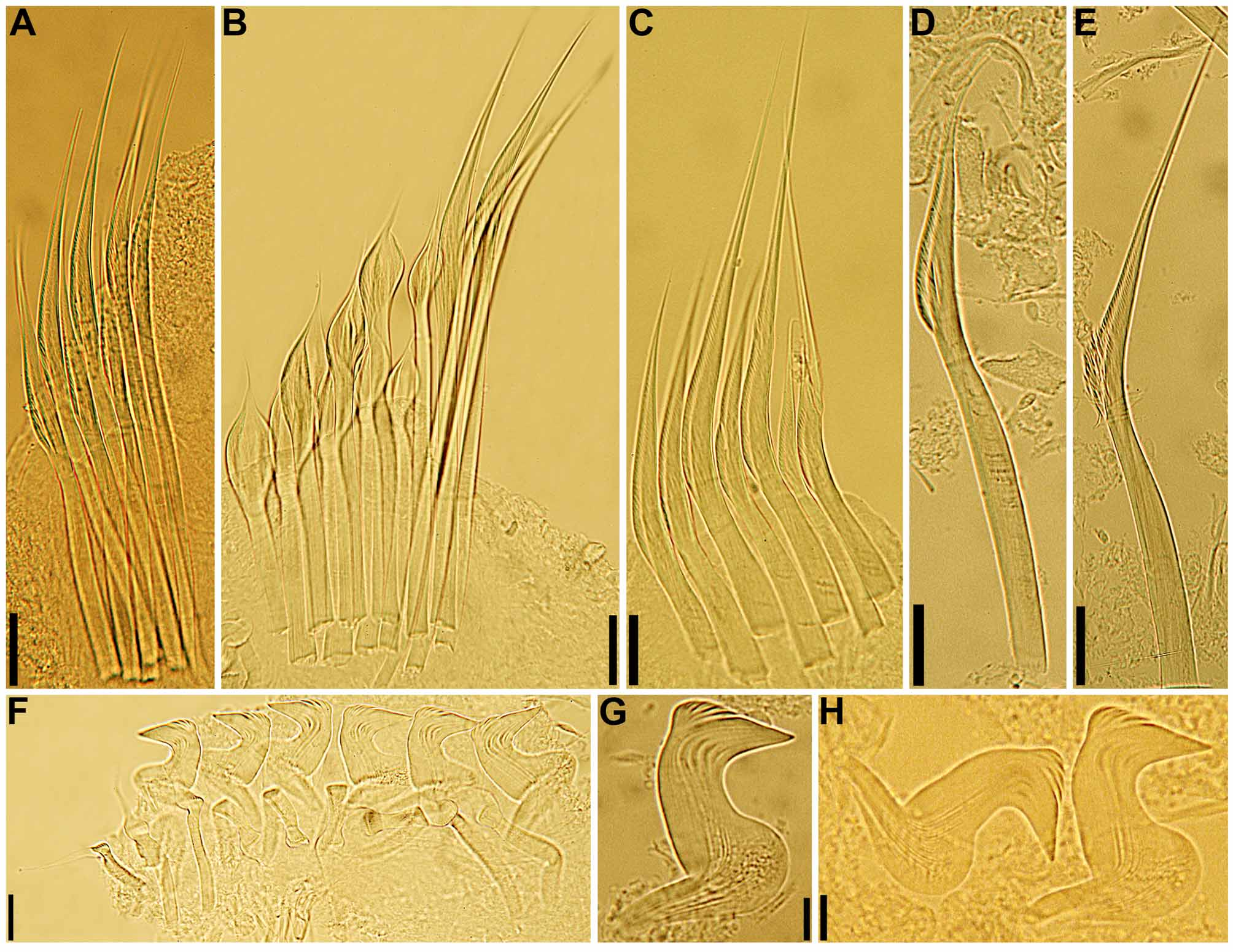
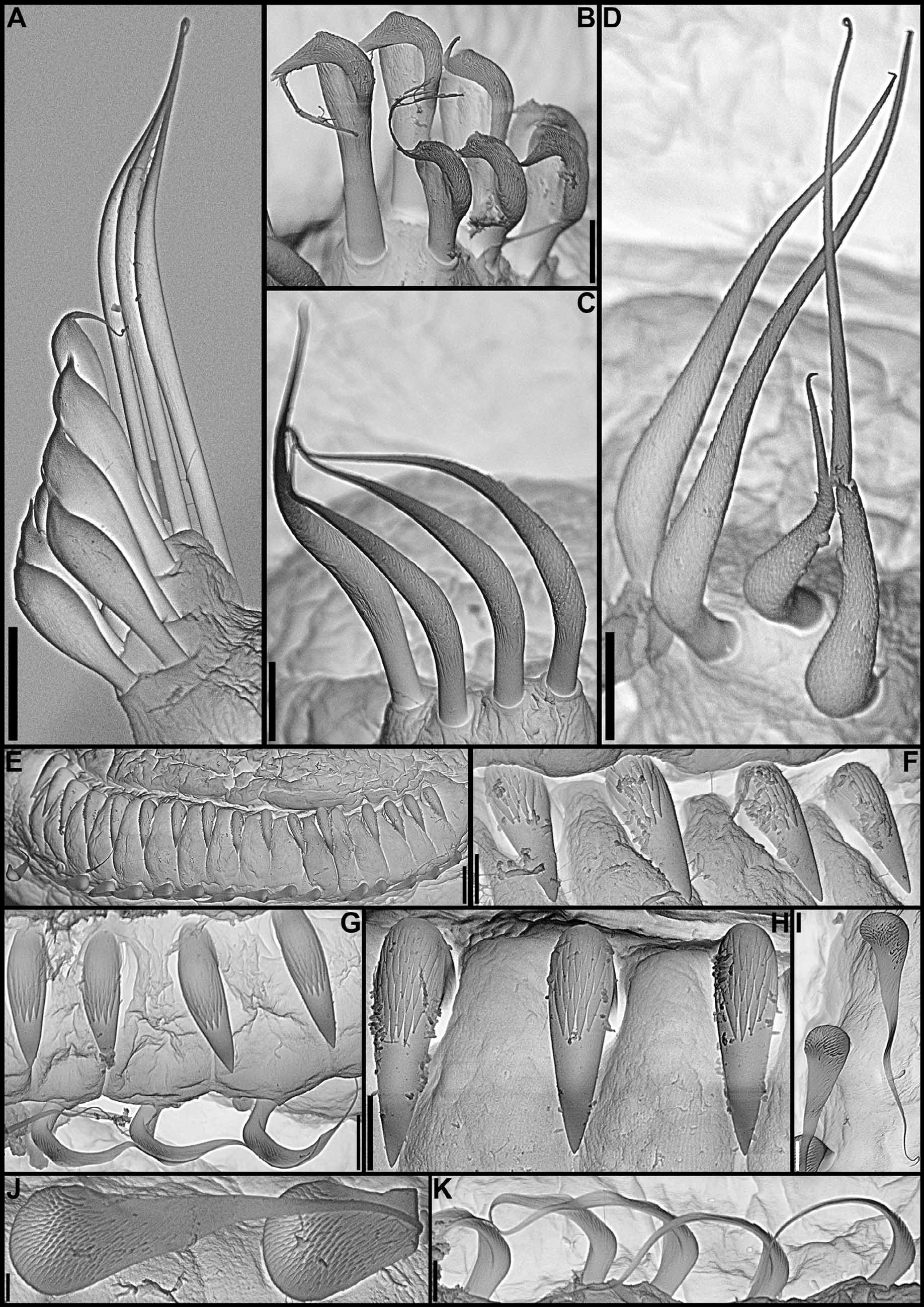
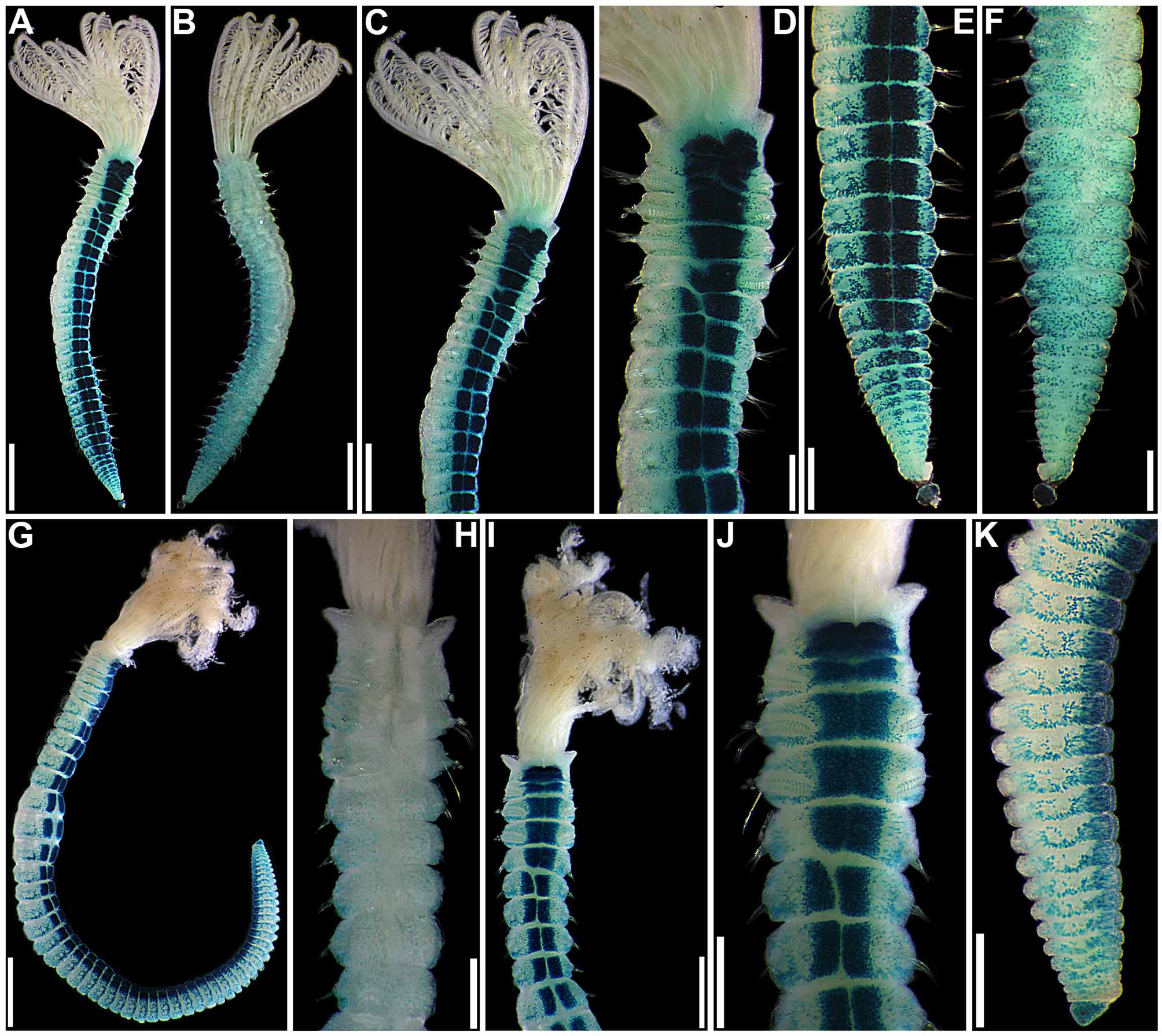
( Figs 7–10 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 , 16 View FIGURE 16 G–K; Table 2)
Material examined. Holotype and 8 paratypes coll. 7 Aug 2003, shallow subtidal (3.05 m) coral-sponge rubble, by G. Hendler, K. Fitzhugh, J. Sanchez, W. Keel, at south side of Cayo Swan (= Isla de Pájaros) (9.453o N 82.3o W), Bahia Almirante, Bocas del Toro Archipelago, Bocas del Toro Province, Caribbean side of Panamá. All type material deposited at the NHMLAC (holotype: LACM-AHF POLY 2404; paratypes: LACM-AHF POLY 2405–2412). Details of each spec. of type series are provided in Table 2.
Additional specimens. USNM 042793: Puerto Rico, Caribbean Sea, 1 spec.; USNM 044906: Key West, Florida, USA, 1 spec.
Comparative material examined. Demonax microphthalmus (CB–T–259–2001; 39027): 1 spec. Slides: radiole; notochaetae, chaetiger 7; uncini, chaetiger 7.
Definition. Two possibilities, depending on character transformation series in the phylogenetic hypothesis involving individuals to which Sabellomma collinae spec. nov., S. minuta , and S. harrisae spec. nov., refers (see Remarks on Specific-level Hypotheses in Sabellomma ): (1) A specific hypothesis, accounting for the absence of dorsal lip pinnular appendages among observed individuals. Dorsal lips without pinnular appendages originated by an unspecified mechanism(s) in a reproductively isolated population of individuals with pinnular appendages almost completely fused to dorsal lips, subsequent to which the new character became fixed in the population by an unspecified mechanism(s), leading to individuals observed in the present, all with dorsal lips lacking pinnular appendages. (2) There are no autapomorphies to provide a basis for the specific hypothesis.
As is discussed below (see Remarks on Specific-level Hypotheses in Sabellomma ), the formal definition of S. collinae as an explanatory hypothesis is tenuous since the explanation of the absence of pinnular appendages is not an autapomorphy in some transformation series.
Description. Measurements. Medium-sized individuals; crown 1–3 mm long, with 4–9 pairs of radioles (Table 2). Complete specimens with trunk 2–10 mm long, 0.2–0.75 mm wide, with 4–5 thoracic, and 18–53 abdominal chaetigers (Table 2).
Pigmentation. Living specimens not observed. Except for radiolar and interramal eyespots, body unpigmented or with very faint brownish pigmentation at base of crown and up to 4–5 radiolar bands. Small interramal eyespots present in all chaetigers, frequently faint and inconspicuous ( Fig. 7 View FIGURE 7 L).
Crown. Radiolar flanges and palmate membrane both absent. Each radiole with four rows of skeletal cells. Radioles with two irregular rows of eyespots along lateral margins; eyespots as single units, each with large lens, more numerous along distal-most two thirds to one-half of radioles, especially in association with transverse pigment bands ( Fig. 7 View FIGURE 7 E–K), less abundant near radiolar tips. Dorsal lips about one-fifth total crown length, basally broad, distally tapering, with radiolar appendages lacking internal skeletal cells; pinnular appendage absent ( Fig. 8 View FIGURE 8 F). Ventral lips low, distally rounded, each continuing ventrally as parallel lamella with large ventral sac above collar ventral lappets ( Figs 7 View FIGURE 7 A, C–G, I–K; 8B–C, E, G–H). Mid-dorsally, branchial lobes with thickened ridges extending along dorsal-most pair of radioles ( Fig. 8 View FIGURE 8 A, D).
Body. Collar fused to body dorso-laterally, well separated from faecal groove, extending anteriorly to level of first pair of parapodia; laterally low, slightly oblique, covering bases of radiolar crown but not origins of radioles; ventrally with one pair of triangular, distally pointed, non-overlapping lappets reaching anterior margin of chaetiger 1, separated from each other by short parallel lamellae; ventrally, between termination of parallel lamellae and anterior margin of first ventral shield, ciliated patch present ( Figs 7 View FIGURE 7 A–K; 8A–E, G–H). Trapezoidal ventral shields, posteriorly indented by neuropodia on thorax, anterior margin of first shield medially indented ( Figs 7 View FIGURE 7 D–E, J–K; 8B–C, G–H). Mid-dorsal faecal groove deep on anterior chaetigers ( Figs 7 View FIGURE 7 B, H; 8A, D). Collar chaetae narrowly hooded, arranged in two rows ( Fig. 9 View FIGURE 9 A); subsequent thoracic notopodia with narrowly hooded superior chaetae, arranged in arc, inferior notochaetae paleate, with long, gently tapering tips, arranged in two rows partially enclosed by superior arc ( Figs 8 View FIGURE 8 A–E; 9B; 10A–B). Thoracic neuropodia with slightly higher than long avicular uncini, crest with 4–5 irregular rows of secondary teeth, covering proximal half of main fang, breast distinct, not extending distally to tip of main fang ( Figs 9 View FIGURE 9 F; 10E, G–H); companion chaetae with rounded, symmetrical, denticulate heads and long, tear drop-shaped, slightly asymmetrical, gently tapering membranes ( Figs 9 View FIGURE 9 F; 10E, G, I–K). Abdominal neuropodia with narrowly hooded chaetae, arranged in two rows, anterior row with shorter chaetae ( Figs 9 View FIGURE 9 C–E; 10C–D). Abdominal uncini similar to thoracic ( Figs 9 View FIGURE 9 G–H; 10F). Triangular, distally rounded pygidium, with 1–3 small, red ocelli at each lateral margin ( Figs 7 View FIGURE 7 M; 8I).
Variation. The specimens of S. collinae sp. nov., examined for the present study presented considerable variation in size, but most other characters did not vary between specimens (Table 2). One specimen (paratype 7), probably a juvenile, has a different number of radioles in right and left halves of the crown. All specimens have the same numbers of thoracic and abdominal noto- and neuropodia on both sides of the body, but there is also slight variation in the number of thoracic chaetigers, the holotype and paratype 7 having 4 thoracic chaetigers, all other specimens having 5 (Table 2). All this may indicate the occurrence of asexual reproduction by scissiparity, as said above for S. minuta .
Etymology. This species is named for Dr. Rachel Collin, director of the Bocas del Toro Marine Station, Smithsonian Tropical Research Institute, Panama, who coordinated the Panamanian survey.
TABLE 2. Morphological variation among the type-series of Sabellomma collinae gen. nov., sp. nov. (variation within parentheses is only shown when there is difference from one side of the body to the other).
continued next page detached chaetae
of
.
more
numerous
eyespots
in most mounted
both
, throughout segments. –; 9 8 A H F; few radiolar eyespots – H G; and abdomen bands, Body devoid. eyespots
data
Additional
,
state
preservation
but
of
crown
the
for
body
extraction
damaged
eyespots
pigmented
,
bands
without
,
third
radioles distal
each
,
from
separated halves
other
fainted and
small
eyespots Interramal
mid-abdominal
anterior and on. stained
Photographed for
Figs
A – 10 K
Crown.
stub
with
SEM on
interramal
conspicuous with
for
.
A
E
Figs
Photographed
8
, –
at two
of beginning
in pieces
with two
. Crown
brownish
stub SEM
half
numerous distal on
of radioles fainted
interramal
small, except for in
good
Complete, from body and 4 short with Crown of on middle third. radioles Crown on SEM stub. conspicuous only methyl green Body Complete mounted, body, eyespots throughout cut Complete, on mounted eyespots more, of pigmentation
,; 2,, 4 13) 1 3:
:
15
Slides segments (: Thorax: notochaetae uncini:, 3; 4 Abdomen neurochaetae, uncini: 13 - -
Distribution. Knight-Jones (1983) assigned to S. minuta specimens with short thorax from Florida (Key West) and Puerto Rico. We have examined those specimens and, although the generic characters of Sabellomma are conspicuous, they do not present dorsal pinnular appendages. Therefore, those specimens are here referred to S. collinae and this species is considered to occur throughout the Caribbean.
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Sabellinae |
|
Genus |