
Discotettix (Discotettix) selysi Bolívar, 1887
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5217.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:86CD1EDF-8C38-4A90-888A-185B8481A6ED |
|
DOI |
https://doi.org/10.5281/zenodo.7409401 |
|
persistent identifier |
https://treatment.plazi.org/id/0129163A-B13B-6002-FCCA-FD6AFEB2F85E |
|
treatment provided by |
Plazi |
|
scientific name |
Discotettix (Discotettix) selysi Bolívar, 1887 |
| status |
|
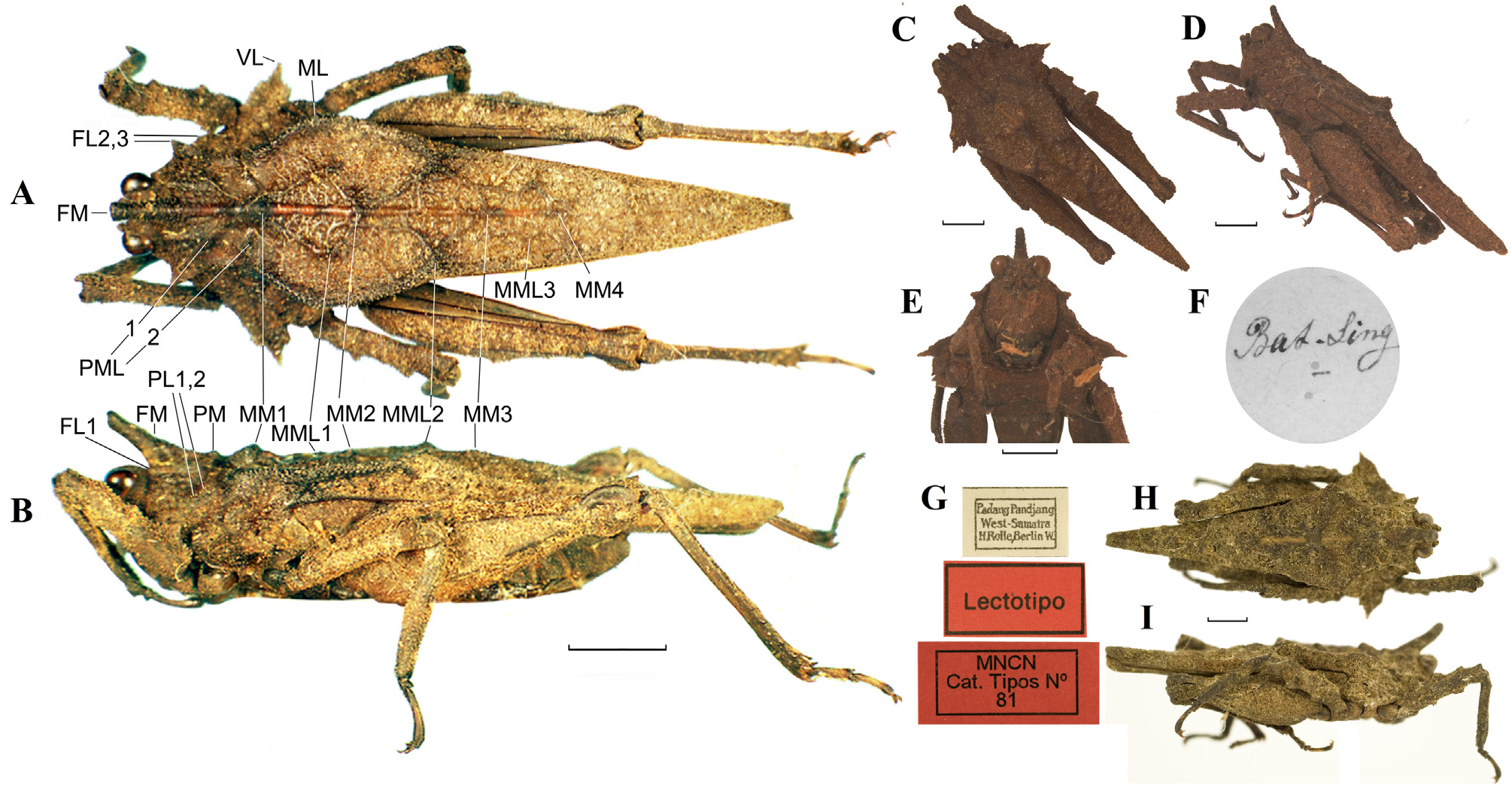
Discotettix (Discotettix) selysi Bolívar, 1887 View in CoL ( Figs 15–17 View FIGURE 15 View FIGURE 16 View FIGURE 17 )
Vernacular name: Sumatran Unicorn Pygmy Devil
Discotettix selysi Bolívar, 1887: 307 View in CoL [original description, type locality: Sumatra]; Hancock 1907a: 6 [listed in catalog]; Kirby 1910: 2 [listed in catalog]; Willemse 1930: 207 [new records], Günther 1938: 301 [partim; new records] Blackith 1992: 46 [listed in catalog], París 1994: 236 [data on the type specimens]; Yin et al. 1996: 866 [listed in the catalog]; Otte 1997: 32 [listed in the catalog].
Discotettix selangori Mahmood et al., 2007: 1276 View in CoL [original description, type locality: Selangor]; syn. n.
Discotettix selangorei : Mahmood et al. 2007: 1275 [lapsus calami].
Acridium (Tetrix) belzebuth (nec Serville): De Haan 1843: 166 [reported for Sumatra, misidentification].
Type locality. Sumatra: Padang Panjang (= originally “Padang Pandjang” on the label) [approximate coordinates 0.45S, 100.416667E].
Note on Discotettix selysi identification history. The species was recorded by De Haan (1843) for the first time under the name D. belzebuth , since only D. belzebuth was described at the time ( D. selysi being described 44 years later). De Haan’s (1843) drawing of D. belzebuth agrees in morphology with D. selysi . The author, in the description, noted the possession of only one well distinctive anterior pronotal projection. Examination of specimens confirmed that De Haan’s records of D. belzebuth from Sumatra belong to D. selysi and there is no evidence of the presence of D. belzebuth on the island.
Material examined.
Type material. SYNTYPES of D. selysi 1♂ Indonesia: Sumatra: Padang Panjang (= on the label “Padang Pandjang”) Collector H. Rolle [the type lacks antennae] ( Fig. 16 View FIGURE 16 ) ( MNCN) ; 1♂ Indonesia: Sumatra 25.XII. [18]84. Soerian ( MHNG) . HOLOTYPE of D. selangori 1♂ Malaysia: Selangor: leg. Brokurtak ( UKM), Sabah Forestry Department, East Malaysia ).
Misidentified museum material. Identified by Günther (1938) as D. selysi : 1♂ Indonesia: Sumatra: west coast, Anai Kloof [500 m a.s.l.] 1926. Collector E. Jacobson, det. K. Günther ( SMTD); (5–6) 2♀♀ Indonesia: Sumatra: Excell. v. Studf., collector and date unknown, det. K. Günther ( MFN); Identified by De Haan (1843) as D. belzebuth : 1♂ + 2♀♀ + 2 nymphs (sex indeterminable) Indonesia: Sumatra: Bat. Sing. [= W Sumatra Isl., Mt. Singgalang volcano] (collector and date not specified in labels), as D. selysi det. J. Skejo et J. Tumbrinck ( Fig. 16 View FIGURE 16 ) ( NCB-RMNH);
Additional museum material. 2♀♀ Indonesia: Sumatra: Maninjau, Puncak, Lawang [600–950 m a.s.l. forest] 17.IV.1995. Collector Sigfrid Ingrisch, det. J. Tumbrinck ( CJT, ZFMK) ; 3♂♂ Indonesia: Sumatra: Mt. Tandikat [600–900 m a.s.l.] VII.2009. Collector Jakl, det. J. Tumbrinck ( ZMUH) ; 1♂ Indonesia: Sumatra: North Sumatra Prov., Roburan Dolok, Panyabungan Selatan, Mandailing , Natal Regency (bamboo stand) 0°44’52.83’’N 99°31’30.68’’E [723 m a.s.l.] 7.X.2019. Collector Fajar Kaprawi, det. J. Tumbrinck ( CJT) GoogleMaps ;
Additional material from online social media. 2 specimens, sex unidentifiable (lateral and dorsal habitus) Malaysia: Peninsular Malaysia: Kuala Lumpur XII.2014. Photographer Pang Way, det. J. Skejo et J. Tumbrinck ( Figs 17 View FIGURE 17 , 18 View FIGURE 18 ) (Facebook) .
Distribution. This species inhabits the rainforests of southern Peninsular Malaysia and of Sumatra, where it can be found on tree bark and roots ( Bolívar 1887; Hancock 1907a; Günther 1938; Mahmood et al. 2007, our data).
Taxonomic notes on Discotettix selangori and D. doriae . The description and the measurements of the recently described Discotettix selangori Mahmood, Idris et Salmah, 2007 (type locality Malaysia: Selangor) completely fit that of D. selysi . The new name for the D. selysi population on the Malaysian peninsula does not provide any new information thus we synonymize D. selangori syn. n. with D. selysi . The authors of the former were not aware of D. selysi morphological variability and distribution. In the description ( Mahmood et al. 2007), the epitheton was “selangori”, while in the key and under the drawing it was written as “selangorei” ( Mahmood et al. 2007). As the first reviewers, according to the ICZN, we pick selangori as the original spelling, while “ selangorei ” is considered a misspelling. We do not agree with Günther’s (1938) synonymy of D. doriae and D. selysi , since a few clear morphological differences can be found (small FM in D. doriae , and swollen antennal segment in D. doriae ) thus we regard D. doriae as a separate species with its own traits and distribution (see a paragraph on D. doriae ).
Diagnosis. The species is morphologically similar to its congeners that do not have strong pronotal projections of the disc, i.e. to D. doriae from Mentawai and D. aruanus from Aru Isl. Discotettix selysi can be distinguished from D. doriae by the following set of characters: (1) D. doriae has FM small and narrow, covering vertex only partially, while in D. selysi FM is large, long, and covering entire vertex); (2) D. doriae is of smaller body size (pronotum length only 16 to 17 mm in females) than D. selysi (pronotum longer than 20 mm in females); (3) D. doriae has shorter and stouter antennae with swollen 6 th, 7 th and 8 th antennal segments, while D. selysi does not have swollen segments and (4) D. selysi has more elongated and slender fore and mid femora, while D. doriae has strong teeth on their ventral and dorsal margins. From D. aruanus sp. n., D. selysi can be distinguished by (I) different shape of the antennae (margins stronger, more saw-like in D. aruanus ); (II) different shape of FM (not exceeding the head in D. selysi , exceeding the head in D. aruanus sp. n.); (III) less elevated and less triangular PM and MM1; (IV) legs more robust and toothed in D. selysi than in D. aruanus sp. n., and (V) by larger body size (pronotum length more than 20 mm in females of D. selysi , while less than 17 mm in females of D. aruanus sp. n.). The species is easily distinguished from D. sumatrensis sp. n. from Sumatra, D. belzebuth from Borneo, D. kirscheyi sp. n. from NE Borneo by the lack of strong spines on the pronotal disc and from the Filipino D. scabridus by numerous characters: (I) bifurcation of the frontal costa between the eyes (on the lower margin of the compound eyes in D. scabridus ), (II) FM high and developed (present as a small tubercle in D. scabridus ), (III) reduced MM, not highly protruded (higher, compressed and saw-like in D. scabridus ), (IV) lateral and humeral carinae are granulated (toothed in D. scabridus ) and (V) larger body size. From D. belzebuth , D. selysi can be easily separated by (I) less widened antennal segments, (II) FM being the only large medial projection (disc of pronotum with many spine-like MM, ML and MML in D. belzebuth ), (III) smoother lateral and humeral carinae (not as equipped and toothed as in D. belzebuth ), (IV) hind femora bearing large lappets, (V) fore and mid femora being more armed and stouter, FM shorter and not decurved as in D. belzebuth . From D. sumatrensis sp. n. the species can be distinguished by (I) completely black antennae, (II) FM being the only large medial projection, pronotum without elevated MM, ML and MML and (III) larger body size. Discotettix kirscheyi sp. n. has (1) smaller FM than D. selysi, (II) larger MM1 and MM2, (II) higher MML2s, (III) more robust fore and mid femora with stronger lobes, (IV) less specialized subapical antennal segments, (V) and is smaller in size. Finally, from D. doriae the species can be separated by (I) longer FM, (II) more slender appearance, (III) not swollen subapical antennal segments, and (IV) larger body size.
Redescription ( Fig. 15 View FIGURE 15 ).
General features. Large-sized and relatively robust species (16–20 mm). Body finely granulated; pronotum slightly rugose, with numerous small tubercles and net-like elevations. The chitinous surfaces are smooth and without tubercles in places, while the rest is strongly granulated. The anterior part of the pronotum bears several strong protuberances, while other projections are reduced ( Fig. 15 View FIGURE 15 ). Macropronotal.
Coloration. The general color dark brown, but may be of brighter tints of brown or even dark greenish. Pronotal carinae and projections darker and of a different color than the rest of the body. Median pronotal carina from dark orange to bright red ( Figs 16 View FIGURE 16 , 17 View FIGURE 17 ). Antennae completely black or dark brown. Maxillary palpi dark brown. The visible part of the tegmen dark brown without any spots. Legs dark brown except for more or less distinct pale rings on tibiae and tarsi. The body usually covered with algae that give a greenish appearance to the specimens. After preservation in alcohol or drying the greenish color disappears.
Head. In dorsal and frontal view, vertex 2.3 times as wide as an eye. Lateral carinae considerably raised and granulated. Fossula deep, but not easily observable because it is covered by large FM of the anterior pronotal margin. Lateral ocelli situated just below the level of the lower margin of a compound eye. Antennal groove significantly below the lower margin of a compound eye. In frontal view, frontal costa bifurcated at the level of lateral ocelli into facial carinae, forming narrow parallel scutellum ( Fig. 15E View FIGURE 15 ). Antennal groove slightly wider than the frontal costa. Antenna 13-segmented (but in male looks like 12-segmented, because the 13 th segment is very small and not visible under an optical microscope, only under SEM): scapus (1 st antennomere) and pedicel (2 nd antennomere) massive; basal segments (3 rd to 6 th) elongated and circular in cross-section; central or subapical segments (7 th and 8 th) strongly widened, pennate, 8 th being the widest antennal segment (about 2.7–2.8 times as long as wide); apical segment 9 th elongated and pennate, smaller than the subapical and much larger than the rest of the apical segments; apical segments 10 th to 13 th reduced, very small, and borders between them barely visible. Antennomeres 3 rd to 9 th bearing saw-like margins, because of the presence of the large basiconic sensilla. When the body and the antennae are covered with algae and moss, the 8 th segment is always free of epizoic organisms.
Pronotum. Pronotum rugose and granulated with numerous small tubercles and net-like elevations, but in some parts smooth and without tubercles (parts of the pronotal disc and some intervals of the median carina of the pronotum). The anterior part of the pronotum bears large FM, and a few medium-sized and small protuberances. The posterior process of the pronotum slender, surpassing hind knees for more than a half-length of the hind femur or less. The disc of the pronotum almost completely flat. A small depression on the disc positioned between the near bases, then the disc becomes slightly elevated again. Caudad, pronotum gradually descending. Disc rich in net-like elevations, more distinct in the places of the interhumeral carinae. The median carina of the pronotum bears unpaired projections of variable size: high and digitate FM, directed upwards and forwards above the head and covering the whole area of the fastigium of the vertex; small triangular PM, a bit larger triangular MM1, small MM2; and completely reduced MM3 and MM4. Prozona subsquare. Prozonal and extralateral carinae in prozona distinct, surpassing anterior margin of pronotum as dentiform small FL1 and larger FL2, FL2. FL3 small, dentiform. Behind the FM medial carina extended along the whole length of pronotum from the anterior margin to the pronotal apex, slightly undulated by the low aforementioned projections. PML1 small, PML2 even smaller. MML1 almost indistinct; MML2 distinct; MML3 very small, while MML4 and MML5 completely indistinct. PL1 and PL2 small. Humeral angle obtuse with pointed apex. ML small, reduced. Pronotum apex narrow, shallowly excised. The lower part of the lateral pronotal lobe with serrate anterior and posterior margins, elongated as spine-like VL, directed strongly outwards and backward ( Fig. 15A, B View FIGURE 15 ).
Wings. The visible part of the tegmen elliptical. Hind wing with scalloped inner margin, a few millimeters shorter than the apex of the pronotal process. In living specimens, the tegmina are covered by photosynthetic microbes (most probably algae and cyanobacteria), so are greenish in appearance.
Legs. Fore and mid femora relatively robust, compressed laterally; with dorsal and ventral margins serrated with genicular tooth on the knees and additional 2–3 strongly projected and sharp teeth on each margin. Hind femur with wrinkled margins, one small protuberance situated on the ventral external carina. Genicular tooth large, while antegenicular tooth almost indistinct. Both sides of the dorsal margin of the hind tibia finely serrated, with 3–4 outer and 1–3 inner larger teeth.
Abdominal apex. Male subgenital plate in lateral view about two times as long as high. Female subgenital plate in ventral view with a longitudinal keel in the median region and with a triangular protrusion in the middle of the posterior margin. Ovipositor robust, dorsal valvae robust, ventral valvae slender; all valvae serrate.
Measurements. BL ♂♂ 14.41–14.88 mm, ♀♀ 16.01–16.28 mm; PnL ♂♂ 17.85–18.35 mm, ♀♀ 20.46–21.11 mm; PnW ♂♂ 8.06–8.14 mm, ♀♀ 9.27–9.48 mm; AnL ♂♂ 7.68– 7.22 mm, ♀♀ 7.18–7.49 mm; TL ♂♂ 2.44–2.51 mm, ♀♀ 2.78–2.91 mm; TW ♂♂ 0.77–0.81 mm, ♀♀ 1.03–1.11 mm; fFL ♂♂ 4.49–4.55 mm, ♀♀ 5.07–5.12 mm; fFW ♂♂ 0.78–0.84 mm, ♀♀ 0.92–0.94 mm; mFL ♂♂ 4.52–4.71 mm, ♀♀ 4.77–5.01 mm; mFW ♂♂ 0.99–1.01 mm, ♀♀ 1.03–1.14 mm; hFL ♂♂ 8.51–8.68 mm, ♀♀ 9.97–10.12 mm; hFW ♂♂ 2.65–2.72 mm, ♀♀ 3.05–3.2 mm; OvL ♀♀ 1.39–1.43 mm; AnL/fFL ♂♂ 1.68–1.7, ♀♀ 1.59–1.69; VW ♂♂ 1.01–1.05 mm, ♀♀ 1.36–1.48 mm; EW ♂♂ 0.39– 0.47 mm, ♀♀ 0.51–0.71 mm; VW/EW ♂♂ 2.23–2.49, ♀♀ 2.43–2.67; SW ♂♂ 0.32–0.36 mm, ♀♀ 0.33–0.42 mm; AgW ♂♂ 0.21–0.33 mm, ♀♀ 0.34–0.39 mm; ScW ♂♂ 0.22–0.27 mm, ♀♀ 0.24–0.29 mm; SW / AgW ♂♂ 1.49–1.53, ♀♀ 1.44–1.58; SW/ScW ♂♂ 1.21–1.45, ♀♀ 1.2–1.34; As–L/W ♂♂ 2.69–2.78, ♀♀ 2.71–2.81; PrzW ♂♂ 3.44–3.52 mm, ♀♀ 4.68–4.71 mm; PrzL ♂♂ 3.93–4.01 mm, ♀♀ 4.74–4.79 mm; Prz–W/L ♂♂ 0.86–0.88, ♀♀ 0.98–1.02; TL/ TW ♂♂ 2.91–3.09, ♀♀ 2.7–2.88; mFW/TW ♂♂ 1.19–1.25, ♀♀ 0.98–1.09; fFL/fFW ♂♂ 5.42–5.61, ♀♀ 5.51–5.71; mFL/mFW ♂♂ 4.32–4.66, ♀♀ 4.46–4.63; hFL/hFW ♂♂ 3.21–3.28, ♀♀ 3.21–3.27; T1L/T3L ♂♂ 1.01–1.08, ♀♀ 1.01–1.02.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Discotettix (Discotettix) selysi Bolívar, 1887
| Skejo, Josip, Pushkar, Taras I., Kasalo, Niko, Pavlović, Marko, Deranja, Maks, Adžić, Karmela, Tan, Ming Kai, Rebrina, Fran, Muhammad, Amira Aqilah, Abdullah, Nurul Ashikin, Japir, Razy, Chung, Arthur Y. C. & Tumbrinck, Josef 2022 |
Discotettix selysi Bolívar, 1887: 307
| Hancock, J. L. 1907: 6 |
| Bolivar, I. 1887: 307 |