
Pseudopaludicola hyleaustralis, Pansonato, André, Morais, Dráusio H., Ávila, Robson W., Kawashita-Ribeiro, Ricardo A., Strüssmann, Christine & Martins, Itamar A., 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.282705 |
|
DOI |
https://doi.org/10.5281/zenodo.6495034 |
|
persistent identifier |
https://treatment.plazi.org/id/03F487E4-FF96-4B02-FF02-F968698BF945 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudopaludicola hyleaustralis |
| status |
sp. nov. |
Pseudopaludicola hyleaustralis sp. nov.
( Figures 2–3 View FIGURE 2 View FIGURE 3 )
Holotype. Adult male ( UFMT 5560; Figure 2 View FIGURE 2 ) collected on 10 January 2007 by A. Pansonato, C. Cavlac, D. H. Morais and E. Brito, on the right margin of the Apiacás River (Salto do Apiacás waterfalls; 10°20'S; 56°56'W), municipality of Alta Floresta, state of Mato Grosso, Brazil.
Paratypes. Eight adult males ( UFMT 5555–9; UFMT 5561–3) and three adult females ( UFMT 5554, UFMT 5564–5) collected between 07–12 January 2007, by the same collectors and at the same locality of the holotype; ten adult males ( UFMT 6056, 6058, 6116, 8267, 8270, 8273, 8275–9) and one juvenile ( UFMT 8288) collected between 5–14 August 2007 by R. W. Ávila and R. A. Kawashita Ribeiro, on the left margin of São João da Barra River (10°18’S; 57°40’W), municipality of Nova Bandeirantes, Mato Grosso, Brazil; three adult males ( UFMT 9197, 9200, 9203), three adult females ( UFMT 9196, 9201–2), and five juveniles ( UFMT 9191–3, 9205, 9207) collected between 16–21 May 2009 by T. Mott, A. Pansonato and A. C. Bortoleto in the federal conservation unit "Estação Ecológica do Rio Roosevelt" (09°06’S; 60°51’W), municipality of Colniza, Mato Grosso, Brazil; one adult male ( UFMT 11328) collected on 27 October 2004 by C. Strüssmann and R. A. Kawashita Ribeiro near the Cachoeira dos Patos (10°12'S; 59°27'W), about 7 km upstream from Dardanelos and Andorinhas falls, municipality of Aripuanã, Mato Grosso, Brazil.
Diagnosis. Pseudopaludicola hyleaustralis sp. nov. is assigned to the genus Pseudopaludicola based on the presence of hypertrophied antebrachial tubercle ( Lynch 1989; Lobo 1995). The species cannot be assigned to the P. pusilla group, due to the absence of T-shaped terminal phalanges ( Lynch 1989). In the genus, Pseudopaludicola hyleaustralis sp. nov. can be considered a medium-sized species, characterized by the combination of the following characters: (1) SVL 12–16 mm in males and 13–17 mm in females, (2) snout rounded both in dorsal and lateral views, (3) upper eyelids smooth, (4) toe tips not expanded laterally, (5) advertisement call composed of non-pulsed notes emitted in series of 11–74 notes, (6) note duration 0.03– 0.05 s, (7) mean dominant frequency of advertisement call 3938±158.65 Hz (3540–4379).
Comparison with other species. Pseudopaludicola hyleaustralis sp. nov. can be distinguished from all congeneric species belonging to the P. pusilla group ( P. boliviana , P . canga, P. ceratophyes , P . llanera, and P. pusilla ) by the absence of either T-shaped terminal phalanges or expanded toe tips (disks or pads). The new species can also be differentiated from selected members of this group by the following characters (characters of compared species in parentheses): from P. boliviana , P. ceratophyes , and P. llanera , by the absence of palpebral tubercles (tubercles evident on upper eyelids; Lynch 1989); from P. ceratophyes , by the presence of toes only basally webbed (toes approximately 1/2 webbed; Rivero & Serna 1984; Lynch 1989); from P. pusilla and P. l l a n e r a, by the snout rounded in dorsal view (snout subacuminate and acuminate, respectively, in dorsal view; Lynch 1989); from P. canga , by the absence of dorsal folds (presence of dorsal folds; Giaretta & Kokubum 2003).
From Pseudopaludicola species not belonging to P. pusilla group ( P. falcipes , P. giarettai , P. m i n e i r a, P. murundu , P. m y s t a c a l i s, P. riopiedadensis , P. saltica , P. serrana , and P. ternetzi ) the new species differs: from P. falcipes , by the presence of an abdominal fold (abdominal fold interrupted or absent; Lobo 1996; Lavilla & Cei 2001); from P. murundu , P. saltica , and P. serrana , by the shorter hindlimbs, with tibio-tarsal articulation reaching the nostrils (presence of very long hindlimbs, with tibio-tarsal articulation reaching beyond the end of the snout; Lobo 1996; Toledo 2010; Toledo et al. 2010); from P. g i a re t t a i, by the smaller body size (16.2–18.0 mm SVL in males) and by the relative length of fingers (finger I shorter than finger IV; Carvalho 2012); from P. mystacalis , by the smaller body size (16.5–18.6 mm SVL in males and 17.3–22.7 mm SVL in females; Haddad & Cardoso 1987); from P. m i n e i r a, by head longer than wide (head as long as wide; Pereira & Nascimento 2004); from P. ternetzi , by the smaller body size (SVL of males 16.0– 18.6 mm; SVL of females 16.0– 22.2 mm; Lobo 1996); from P.
riopiedadensis , by finger I shorter than II, tibio-tarsal articulation reaching the nostrils, and head longer than wide (finger I equal to finger II, tibio-tarsal articulation reaching the eyes, and head wider than long; Mercadal de Barrio & Barrio 1994).
Advertisement call structure also distinguish Pseudopaludicola hyleaustralis sp. nov. from all other nine congeners for which call descriptions are available ( P. boliviana , P. c a n g a, P. falcipes , P. giarettai , P. m in e ir a, P. murundu , P. m y s t a c a l i s, P. saltica , and P. serrana ). The call of the new species differs from the calls of P. b o l i v i a n a, P. falcipes , P. m i n e i r a, P. m u r u n d u, P. m y s t a c a l i s, P. saltica , and P. serrana by non-pulsed notes (pulsed notes; Duré et al. 2004; Haddad & Cardoso 1987; Pereira & Nascimento 2004; Toledo 2010; Toledo et al. 2010). Additionally, frequency range is lower in the call of the new species than in the calls of P. falcipes (4200–5800 Hz; Haddad & Cardoso 1987), P. m i n e i r a (4306–4823 Hz; Pereira & Nascimento 2004), P. m u r u nd u (4710–7250 Hz; Toledo 2010), P. saltica (5000–6600 Hz; Haddad & Cardoso 1987), and P. serrana (3120–8490 Hz; Toledo 2010), and slightly higher than in the call of P. mystacalis (2800–4100 Hz; Haddad & Cardoso 1987). From P. b o l i v i a n a, the advertisement call of the new species also differs in the higher number of notes (only five notes; Duré et al. 2004). Amongst species for which non-pulsed notes were described, the call of Pseudopaludicola hyleaustralis sp. nov. differs from that of P. c a n g a by a higher number of notes (3–19; reanalysis of data provided by Giaretta & Kokubum 2003), higher call series duration and lower note repetition rate (call series duration 0.2– 0.5 s and 690–944 notes per minute; reanalysis of data provided by Giaretta & Kokubum 2003). From P. giarettai , the new species differs by call series composed of short notes and by higher note repetition rate (isolated and long notes, and 117–153 notes per minute; Carvalho 2012).
Description of holotype. Snout rounded both in dorsal and lateral views; head longer than wide; tympanum indistinct; nostrils slightly protuberant, directed anterolaterally; pupil horizontally elliptical; vomerine teeth absent; tongue ovoid, covering the entire floor of mouth and posteriorly free; vocal sac single, subgular. Surfaces of dorsum, venter, upper eyelids, belly and throat smooth. Arms robust, with two subconical antebrachial tubercles, the distal one scarcely perceptible; fingers free, relative lengths IV<I<II<III; inner metacarpal tubercle oval; outer metacarpal tubercle rounded; subarticular finger tubercles enlarged, rounded; thumb with a fine nuptial asperity on the base; supernumerary finger tubercles small, rounded. Hindlimbs robust, long; thigh longer than shank, foot longer than thigh; a well developed dermal fold from the inner metatarsal tubercle to mid length of tarsus; thickening of the distal end of this tarsal fold resembles a tubercle; toes basally webbed, relative lengths I<II<III<V<IV; inner metatarsal tubercle oval; outer metatarsal tubercle rounded; subarticular toe tubercles enlarged, rounded, supernumerary toe tubercles absent.
Color. In life ( Figure 3 View FIGURE 3 A–B), dorsum and limbs are brown, with either green and brown spots or stripes. Belly white; ventral view of legs yellowish white and vocal sac beige with dark dots; iris reddish-brown superiorly and grayish-brown inferiorly; upper lip with vertical wide green stripes (see Figure 3 View FIGURE 3 ). In preservative, dorsum and dorsal surface of limbs grayish brown with dark brown spots or stripes; belly and ventral surface of legs light beige; vocal sac grayish.
Variation. Sexual size dimorphism is apparent, with females attaining larger size than males ( Table 1). Adult males have a beige vocal sac, either unpigmented (in 55.8% of the 29 examined males) or with grayish vermiculations (44.2%). Dorsal coloration is highly variable, from light brown to dark gray, with nearly one third of the specimens (35.2% of the 35 examined specimens) presenting a pair of lateral light stripes from the eyes to the groin. Ventral coloration is light beige, either unpigmented (55.8% of the examined specimens) to heavily pigmented (26.4%), with some specimens (17.8%) presenting an unpigmented light beige belly but pigmented vocal sacs and ventral surfaces of thighs. Only one specimen out of 35 (UFMT 8275) has a thin vertebral stripe from the tip of the snout to vent.
Pseudopaludicola hyleaustralis Pseudopaludicola canga
Marabá Curionópolis Male paratypes Female paratypes Males Females Males Holotype (N=29) (N=6) (N=6) (N=5) (N=6)
Snout-vent length 15.9 14.3±1.1 15.4±1.7 15.2±0.63 18.2±1.85 15.7±0.6 (12.2–16.1) (13–17.5) (14.6- (15.5-20.1) (14.6-16.5) 16.2)
Head width 5.3 4.8±0.5 5.3±0.5 5.4±0.53 6.1±0.52 5.3±0.2 (3.7–5.8) (4.4–5.7) (4.6-6.0) (5.4-6.6) (5–5.6)
Head length 6.1 5.3±0.5 5.7±0.8 4.8±0.24 5.3±0.36 6.3±0.3 (4.5–6.1) (4.6–6.5) (4.5-5.1) (4.9-5.9) (5.8–6.8)
Tibia length 6.7 6.9±0.6 7.1±0.5 7.6±0.46 8.3±0.78 7.7±0.3 (5.8–8.0) (6.6–7.9) (7.1-8.3) (7.1-9.3) (7.3–8.0)
Thigh length 7.1 6.8±0.5 7.4±0.6 7.9±0.3 8.9±0.58 7.1±0.4 (5.9–7.7) (6.8–8.5) (7.4-8.2) (8.3-9.5) (6.5-7.5)
Foot length 8.4 7.8±0.6 8.1±0.3 8.6±0.51 9.2±0.72 8.4±0.4 (6.8–8.9) (7.1–8.5) (7.8-9.1) (8.1-9.9) (7.9-9.0)
Hand length 4.2 3.9±0.4 4.1±0.3 4.4±0.38 4.5±0.5 4.0±0.2 (3.3–4.7) (3.7–4.5) (3.7-4.7) (4.0-5.2) (3.8-4.4)
Eye diameter 2 1.9±0.2 1.9±0.2 2.2±0.22 2.2±0.24 1.8±0.1 (1.5–2.3) (1.7–2.1) (2.0-2.6) (2.0-2.5) (1.8-1.9)
Eye–nostril 1.3 1.4±0.2 1.3±0.1 1.3±0.14 1.5±0.12 1.7±0.1
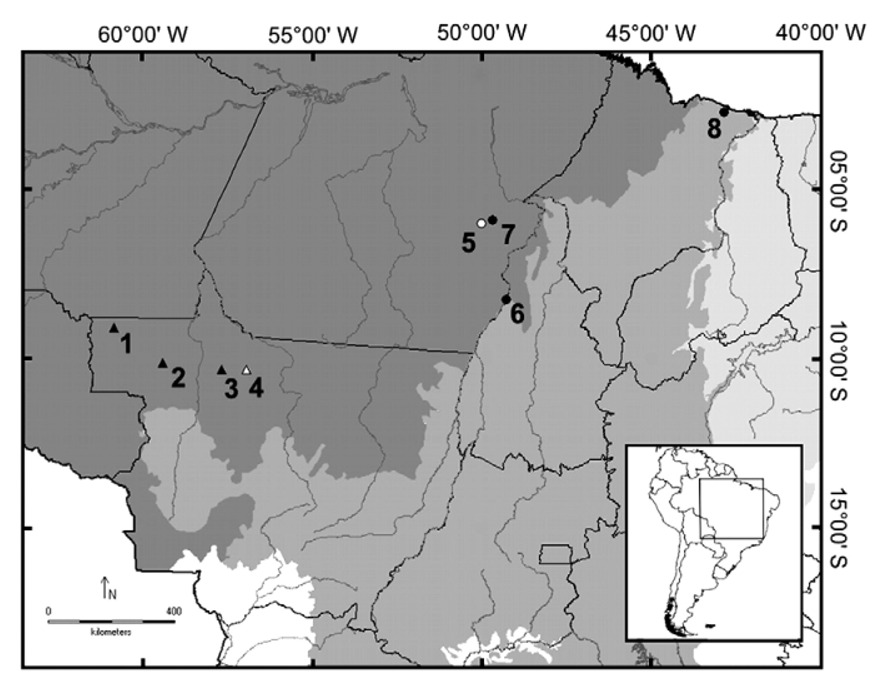
distance (1.2–1.8) (1.1–1.4) (1.1-1.4) (1.4-1.7) (1.6-1.9) Geographic distribution. Pseudopaludicola hyleaustralis is known from four localities in the northern state of Mato Grosso, Brazil (municipalities of Colniza, Alta Floresta, Nova Bandeirantes, and Aripuanã), along the drainages of the Apiacás, Roosevelt and Aripuanã rivers ( Figure 1 View FIGURE 1 ).
Natural history notes. Males of Pseudopaludicola hyleaustralis at Alta Floresta and Nova Bandeirantes were found calling in the evening, until 20:00 h. Males call from the ground, in shallow water with sandy or muddy substrates, in open areas surrounded by primary forest. The single specimen from Aripuanã was found during the day, on the muddy bed of a tiny intermittent tributary of the right bank of the Aripuanã River. At Alta Floresta, couples in axillary amplexus ( Figure 3 View FIGURE 3 B) were observed in January, during the peak of the rainy season.
Conservation. In most of the localities aforementioned, populations of Pseudopaludicola hyleaustralis are under strong pressure due to habitat destruction. In the municipalities of Alta Floresta, Aripuanã, and Nova Bandeirantes, the main causes of habitat loss are due to the construction of hydroelectric power plants, selective logging, and mineral exploration. The situation seems to be better at Colniza, where some populations might be protected in the 800 km 2 of the federal conservation unit "Estação Ecológica do Rio Roosevelt".
Description of the advertisement call. We analyzed 135 calls of two specimens of Pseudopaludicola hyleaustralis (holotype UFMT 5560; paratype UFMT 5561), both recorded at Alta Floresta, Mato Grosso, Brazil. A typical advertisement call of the new species consists of call series composed of 11–74 (43.3±18.7) non-pulsed notes (Figure 4), with a mean call series duration of 4.25± 2.2 s (range 1.14–8.15), emitted at regular intervals of 46.78± 11.63 s (29.73–58.79). Mean duration of each note is 0.03± 0.003 s (0.025–0.049). Each note is followed by internote intervals of 0.078± 0.022 s (0.035–0.171) and the average rate of notes per minute is 561.45±44.5 notes/ min (503.76–623.21). Mean frequency ranges from 3540–4379 Hz, and mean dominant frequency is 3938±158.65 Hz (3605–4165.3). Each note has a slight ascendant frequency modulation in its first half, and another ascendant frequency modulation in its last half.
Etymology. The specific epithet, an adjective, is derived from the Greek word “ Hylea ” meaning “great forest”, and the Latin word australis, meaning “south” or “southern”, which together is translated as “from the southern part of the Hylea". Hylea was first applied to the Amazon Valley region in the early 19th century by the German scientific explorer Alexander von Humboldt.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.