
Monocoryne bracteata ( Fraser, 1943 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.46301 |
|
DOI |
https://doi.org/10.5281/zenodo.6063907 |
|
persistent identifier |
https://treatment.plazi.org/id/C26087FD-6F0D-E90F-B5AA-FBF37C99FEAB |
|
treatment provided by |
Plazi |
|
scientific name |
Monocoryne bracteata ( Fraser, 1943 ) |
| status |
|
Monocoryne bracteata ( Fraser, 1943) View in CoL View at ENA
Figs 1-3 View Fig. 1 View Fig. 2 View Fig. 3
Symplectanea bracteata Fraser, 1943: 78 View in CoL , pl. 13 fig. 1.
Monocoryne bracteata View in CoL . – Rees, 1958: 17, fig. a-b. – in part Stepanjants et al., 2003: figs 2A & 2C. ‒ Brinckmann-Voss & Lindner, 2008: 1634, fig. 2A-B.
Material examined:
MHNG-INVE-92014, field number Kam05; 15 specimens in ethanol (males and females) and serial histological sections of middle region of males and females; Russia, Kamchatka Peninsula, Avacha Bay, Starichkov Island, 52.7823°N 158.61575°E, 16 m depth, temperature 2°C; collection date 18.06.2014; DNA extracted from 2 specimens preserved in ethanol, DNA isolates 1123 and 1124 gave both the same 16 sequence LN898142 View Materials .
MHNG-INVE-92015, field number Kam06; 1 male and 1 female specimen in ethanol; Russia, Kamchatka Peninsula, Starichkov Island, 52.77915°N 158.61268°E, 7 m depth, temperature 10 °C; collection date 24.06.2009.
MHNG-INVE-92016, field number Kam07; 9 specimens in ethanol, males and females; Russia, Kamchatka Peninsula, Starichkov Island, 52.7796°N 158.6111°E, 10 m depth, temperature 5°C; collection date 17.07.2009; DNA extracted from 2 specimens preserved in ethanol, DNA isolates 1125 and 1126 gave both the same 16S sequence LN898143 View Materials .
ZIRAS; several specimens in ethanol, one with branched polyp body; Russia, Kamchatka Peninsula, Avacha Bay, Starichkov Island, on pebbles, 52.7790°N, 158.6124°E, 8 m depth, temperature 4°C; collection date 0 2.06.2011.
ROMIZ B3646, 1 fragmented specimen with female sporosacs in ethanol; Russia, Okhotsk Sea, Urup Island, 46°N 149°E, 300 m depth; collection date 07.08.1987; elongate sporosacs with 0- 20 egg like bodies.
Type locality: Albatross Station 4253; 57.67194°N 133.67944°W, Thistle Ledge, Stephens Pass, southeastern Alaska, water depth 240-344 m ( Brinckmann-Voss & Lindner, 2008).
Diagnosis: Solitary species of Monocoryne , with or without anchoring filaments, up to 200 widely spaced tentacle-groups comprising 3-10 tentacles (mostly around 6), tentacles grouped in linear arrays. Individuals gonochoristic, gonophores developing in upper axis of tentacle-groups, occurring as sporosacs without canal systems, elongate when fully mature, up to 3-4 times as long as wide, mature females with 3-20 maturing oocytes.
Description of Kamchatka material:
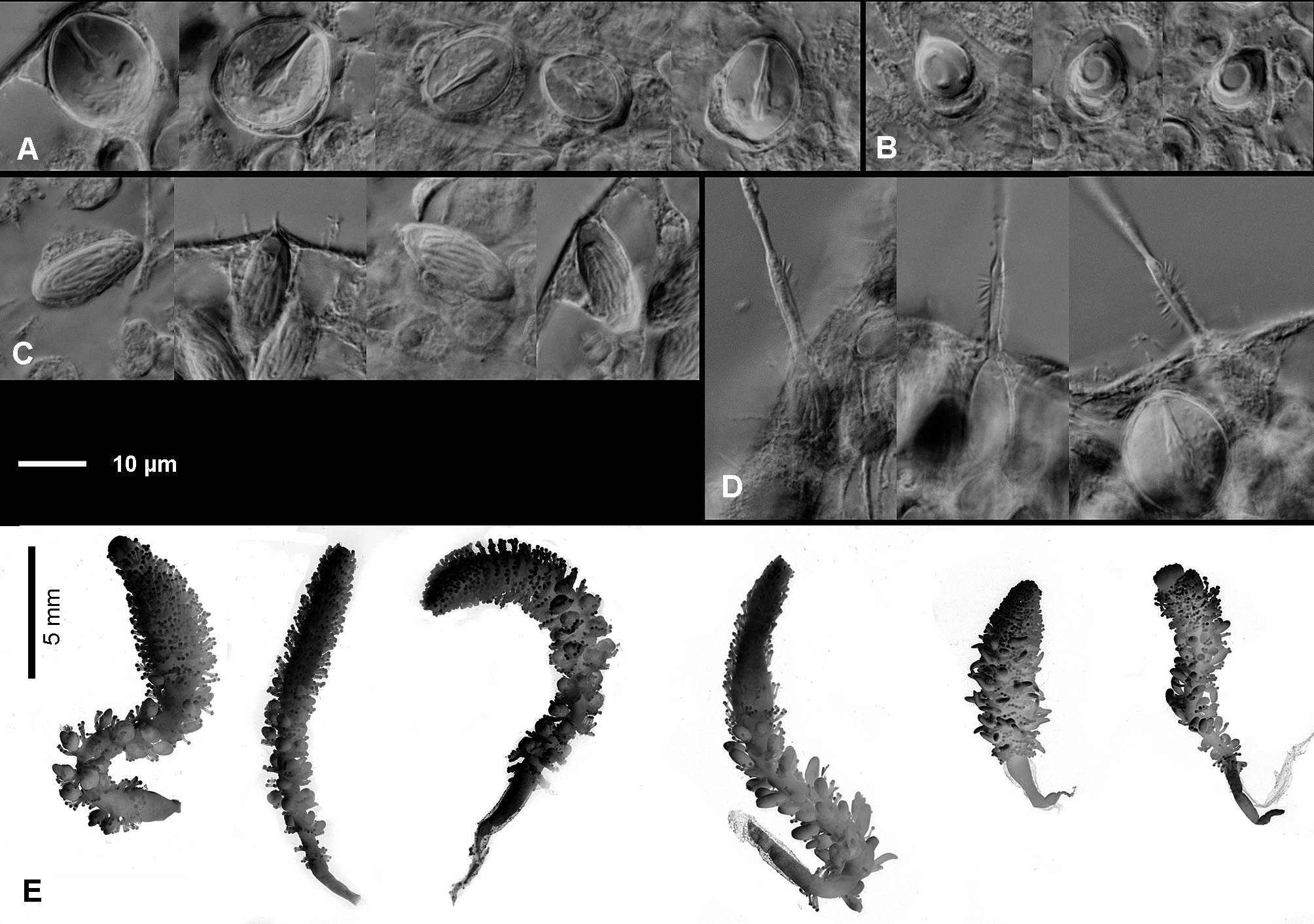
Morphology: Hydroid polyps solitary, vermiform, able to contract to 1/10 of length, not gregarious, subdivided into a basal, tentacle-free foot and an upper, tentaclebearing part. Foot conical, length about 1/6 or less of total height when expanded, up to 1/3 when contracted or preserved, sheathed in an adhering, very thin, filmy perisarc, hardly visible in living animals, loose and wrinkled in preserved specimens. Basal tip of perisarc attached directly to substrate (small stones, shell debris); anchoring filaments absent. Tentacle-bearing zone thin, vermiform, squirming, beset with widely spaced groups of tentacles (in relaxed body). Tentacles capitate, somewhat contractile, up to 200 or more, in groups of 3-10 (mostly around 6), those of a cluster in a single row with bases fused through a common epidermis ( Fig. 2 View Fig. 2 G), rows of tentacles oriented transversely or obliquely to polyp axis. Terminal knobs (capitula) of tentacles spherical, about twice diameter of stalk; tentacle stalks relatively long and thin when extended. Terminal region of polyp with small, shallow hypostome surrounded by solitary tentacles.
Gonophores sporosacs developing singly in upper axils of tentacle-groups, usually confined to lower half of tentacle-bearing part. Sporosacs with distinct spadix, without radial canals or other vestiges of medusa stage ( Fig. 2 View Fig. 2 E-F). Young sporosacs spherical, later becoming elongated, contractile. Fully developed, relaxed male sporosacs oblong, about three times as long as thick ( Fig. 2 View Fig. 2 B). Female sporosacs spherical to spindle shaped when mature ( Fig. 2 View Fig. 2 C-D), containing 1-4 white, egg-like bodies (mature oocytes or clusters of developing oocytes and their nourishing cells). Individual polyps apparently either male or female, never hermaphroditic.
Nematocysts (preserved tissue): Larger stenoteles (10.5- 13)x(14-15)μm; smaller stenoteles (8-9)x(11.5-13)μm; desmonemes (5-6.5)x(7-9)μm; microbasic heteronemes (5-6.5)x(13.5-18)μm ( Fig. 3 View Fig. 3 A-D).
Dimensions: Fully grown expanded polyp 40 mm and more, diameter ca. 0.8 mm; preserved specimens about 10-15 mm. Expanded tentacles up to 1.5 mm long. Sporosacs 0.8-2.1 mm long. Eggs maximally 0.35- 0.5 mm in in size.
Colours: Variable, yellowish-orange ( Fig. 1 View Fig. 1 A-B), pale-pink-orange, to orange-red. Surface of sporosac sometimes with rusty-red pigment clusters ( Fig. 1 View Fig. 1 B). Spadices of sporosacs when fully grown dark brown-red, eggs white when fully mature ( Fig. 2 View Fig. 2 C-D).
Other observation: One polyp with a bifid, Y-shaped body was found.
Biology: Found in depths of 7-16 m in gravel, attached to small stones, shell debris, or tubes of polychaetes.
Distribution: North Pacific; South-eastern Alaska ( Fraser, 1943), Kamchatka Peninsula and Urup Island (this study), depth range 7- 344 m.
Remarks: Hydroids of Monocoryne Broch, 1910 , rare and seldom observed, are rather distinctive in having a large, worm-like body with numerous tentacles that are usually clustered in linear groups ( Fig. 1 View Fig. 1 A). Currently comprising four species ( Schuchert, 2015), the genus has recently been reviewed by Stepanjants et al. (2003). Since that review, a new species ( Monocoryne colonialis ) has been described by Brinckmann-Voss & Lindner (2008) that had previously been mistaken by Stepanjants et al. for M. bracteata . One species of the genus, M. minor Millard, 1966 , is restricted to South Africa. Meanwhile, M. colonialis is unusual in forming small colonies and in having tentacles that are not in groups. These two species can be excluded from consideration in identification of the present material. The two remaining species, the Atlantic M. gigantea ( Bonnevie, 1899) and the Pacific M. bracteata ( Fraser, 1943) appear to be very similar morphologically. After examining type material of both species, Rees (1958) concluded that they could be conspecific. He kept them apart only because the type material of M. bracteata was in such a poor condition that it precluded any appropriate comparison. Both species are rare, M. bracteata was so far only known from type material consisting of two fragmented polyps. Apart from their geographic origin (Atlantic- versus Pacific Ocean), the two are currently only distinguished based on the maximal number of tentacles per cluster (“bract-like” clusters of Fraser, 1943), with 3-4 in M. gigantea and 4-7 in M. bracteata ( Rees, 1956, 1958; Brinckmann-Voss & Lindner, 2008).
Rees (1958) claimed that M. bracteata might have hermaphroditic sporosacs as in M. gigantea (see Johannesen, 1924). However, he did not make histological sections and without them, sex determination in preserved hydroids is difficult. Preserved Kamchatka samples were impossible to sex unambiguously without histological sections. Photos of the type specimen of M. bracteata show sporosacs with a few (4-5) egg-like bodies only, although Brinckmann-Voss & Lindner (2008) depict 7 in their drawing. Fraser (1943) shows many more, but his drawings tend to be somewhat inaccurate and his text states that they are “not numerous”. The sample from Urup Island likewise had sporosacs with spindle-shaped spadices surrounded by 8-20 egg-like bodies (max. size 0.3 mm). However, a few sporosacs had no eggs at all and had presumably spawned already. It is therefore assumed that the low egg numbers per sporosac observed in the Kamtchaka material ( Fig. 2 View Fig. 2 C-D) is due to older sporosacs that had already liberated eggs. What appears as eggs in female sporosacs are often only clusters of nourishing cells surrounding a developing oocyte ( Fig. 2 View Fig. 2 F). Egg numbers per sporosac in M. gigantea are not known, but illustrations in Johannesen (1924) suggest low numbers as in M. bracteata .
The histological examinations ( Fig. 2 View Fig. 2 E-F) showed that the Kamchatka animals are clearly dioecious, in contradistinction to M. gigantea which is hermaphroditic (see Johannesen, 1924). This should be taken as a species specific difference.
As for the cnidome, the heteronemes ( Fig. 3 View Fig. 3 D) might be interpreted as microbasic euryteles with a faint swelling, but as is often the case in studies of preserved material they could be mastigophores instead.
The four 16S sequences obtained from M. bracteata were all either very similar or identical, adding further evidence that all the samples belong to the same species. The sequences are rather distant to all other Hydrozoa 16S sequences, but similar to an unpublished 16S sequence of Monocoryne colonialis (results not shown). To conclude, the numerous polyps from the Kamchatka Peninsula examined here most likely belong to M. bracteata , despite the following differences from the type specimens:
- shallow water (7-16 m) versus deep-water (> 240 m) - no attachment filaments; shorter foot region (2/3 of total height in type)
- fewer eggs per sporosac
- shape of fully grown sporosac more elongated.
The deep water specimen from Urup Island more closely resembles the type material of M. bracteata . All these differences were interpreted here as a combination of intraspecific, environmentally induced, and age-related variations.


| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Monocoryne bracteata ( Fraser, 1943 )
| Peter Schuchert, Nadya Sanamyan & Karen Sanamyan 2016 |
Monocoryne bracteata
| Brinckmann Voss A. & Lindner A. 2008: 1634 |
| Rees W. J. 1958: 17 |
Symplectanea bracteata
| Fraser C. M. 1943: 78 |