
Microporella harmeri Hayward, 1988
|
publication ID |
https://doi.org/ 10.5281/zenodo.207232 |
|
DOI |
https://doi.org/10.5281/zenodo.6184658 |
|
persistent identifier |
https://treatment.plazi.org/id/0387F320-FFE2-FFE5-D4AF-F92FFA8CFC3B |
|
treatment provided by |
Plazi |
|
scientific name |
Microporella harmeri Hayward, 1988 |
| status |
|
Microporella harmeri Hayward, 1988 View in CoL
( Figs 8 View FIGURE 8. M A–F, 9A–D, 10A–F, 11A–D, Table 5 View TABLE 5 )
Microporella harmeri Hayward, 1988 View in CoL , p. 324, pl. 10e; Scholz et al. 2001, p. 222, fig. 5c;? Gordon et al. 2007, p. 51, fig. 3c.
Microporella orientalis View in CoL var. Harmer, 1957, p. 963, pl. 62, fig. 30;?d’Hondt 1988, p. 194.
Microporella orientalis: Aristegui 1984 View in CoL , p. 329, figs 67d–f, pl. 24, figs 7–8, pl. 25, figs 1–2;? Zabala & Maluquer 1988, p. 140, fig. 338; Ryland & Hayward 1992, p. 279, figs 25e–f.
Microporella ciliata View in CoL var. personata: Thornely, 1905 , p. 112.
Material examined. Lebanon: Selaata, 3–8 m, on cave wall, 18–23 October 1999. Red Sea: (1) South Sinai, Ras Mohammed, ‘Yolanda’ wreck, 18 m, on aluminium sheet, 15 May 1983; (2) Safaga Bay, south to Ras Abu Soma, depth 1–20 m, 3 colonies on coral pieces, September 1992; Safaga Bay, west part of Safaga Island, transect A5, 1– 2 m, one colony on Cymodocea sp., September 1992; Safaga Bay, station A 1-2/2, 10 m, sand, one fragment, 27 April 1986; Safaga Bay, transect B2, 42 m, one fragment on bivalve shell, date unknown. Indian Ocean: (1) south Oman, Salalah, near Mirbat, Kelp Bay, right side, depth 9 m, 5 colonies on bivalve shells and coral pieces, 16 January 2009; (2) south Oman, Salalah, near Mirbat, depth 8 m, one colony on bivalve shell, 18 January 2009; (3) Maldive Islands, North Male Atoll, Helengeli Is., March 1983, one colony on bivalve shell, coll. F.F. Steininger; (4) South Africa, Sodwana Bay, 2 Mile Reef, 1993, one colony on coral, coll. B. Riegl. Other material examined: (1) Balavoine Collection, MNHN n 7803, one colony labelled as Microporella umbracula ( Audouin, 1826) , Gulf of Suez, Al Sayad station X. (2) Specimen of Microporella ciliata var. personata from Ceylon, Thornely Collection, NHM n 1999.7.19.18, SEM photos (courtesy M. Spencer Jones). (3) Specimen of M. harmeri from Mauritius, Hayward collection, NHM n 1987.1.18.23, SEM photos (courtesy P.J. Hayward). (4) Specimens attributed to M. orientalis from Heron Island, Stn 25, Ryland & Hayward collection, NHM n 1996.2.23.92, SEM photos (courtesy P.D. Taylor).
Description. Colony small or medium-sized, unilaminar. Autozooids quadrangular, hexagonal or oval, longer than broad (mean L/W = 1.2 in Lebanon, 1.3 in Safaga Bay, 1.3–1.4 in Oman); frontal shield weakly convex, ornamented with rounded grains, perforated by numerous pseudopores (45–90) and 5–9 larger, elongated or oval marginal pores. Ascopore placed at distance from orifice, proximal border equivalent to orifice length or a little smaller, crescentic, median process rounded, either well-developed and broad or small, leaving C-shaped lumen partially filled with sharp denticles, sometimes ramified and fusing in transverse bars, surrounded by low rim. Primary orifice broader than long, with distal edge smooth and proximal edge serrated with 11–20 (11–15 in Selaata, 11–17 in Oman, 16–20 in South Africa) sharp, triangular or conical denticles that are often slightly shorter in middle part of edge. Oral spines thin, vanishing in older ontogeny, 3 in most cases, occasionally 2 or 5. Avicularium single on left or right side, exceptionally paired (1 example in a South African colony), directed distolaterally, located slightly proximal to ascopore, but sometimes in a distal position; opesia notably broad (width of crossbar equal to or a little smaller than width of orifice proximal edge), crossbar complete and robust; rostrum short, channelled, with parallel sides or slightly flared at tip; mandible lanceolate with asymmetrical leaf-like projection, internal side broader than external one; two pointed lateral processes curved basally, leaning against rostrum tip. Double-walled ooecium with membranous ectooecium and calcified entooecium; latter globular, slightly broader than long, granular, bearing ‘pseudopores’ that are distinctly smaller than those of frontal shield, scarcer on proximal third of ooecium; proximal edge forming slightly arched smooth rim; proximal part of ovicell personate, as a raised, granular collar distally adjacent to ascopore, overhanging orifice and fusing with proximal rim of entooecium; this proximal collar sometimes only partially developed or totally absent (2 cases in a South African colony). Ancestrula tatiform with narrow and smooth cryptocyst, bordered by 11 spines.
Lebanon S Sinai Safaga Oman Maldives S Africa Remarks. Microporella harmeri was introduced by Hayward (1988) for specimens from Mauritius characterized in particular by three ephemeral spines, a primary orifice with finely crenulate proximal border, maternal zooids with a peristome formed by a prominent proximal lip, and unpaired avicularia with a large opesia and a lanceolate mandible with broad, asymmetric, lateral flaps. This very particular mandible shape was noticed first by Harmer (1957, p. 504) in specimens from the Indian Ocean recorded as M. orientalis var., synonymized with M. harmeri by Hayward (1988). Microporella orientalis var. was recorded from Eilat by d’Hondt (1988) without comment or illustration. When recording Microporella ciliata var. personata, Busk from the Gulf of Manaar, Ceylon, Thornely (1905, p. 112) specified that "The avicularia on the Ceylon specimens have wing-like membranous extensions of the sides of the mandibles". This record was included by Harmer (1957) in the synonymy of Microporella orientalis var. SEM photos of one of these Ceylon specimens in the NHM (courtesy of M. Spencer Jones) allowed confirmation of its identity with M. harmeri based on the typical personate ovicells, avicularia with particularly broad opesiae and the proximal edge of the orifices having pointed teeth.
Specimens from the Indian Ocean (south Oman, Maldives, South Africa), Red Sea (Ras Mohammed, Safaga Bay) and southeastern Mediterranean ( Lebanon) are ascribed to M. harmeri with a good level of confidence. All of the specimens found on artificial substrata (Ras Mohammed) or on bivalve shells and coral rubble (remainder of material) present orifices with a finely denticulate proximal edge, three thin ephemeral spines, personate ovicells with a raised peristome and single avicularia with large opesia and leaf-like mandibles with asymmetric lobes. Although the number of teeth on the proximal edge of the orifice can vary among these specimens, their shape is remarkably constant ( Fig. 10 View FIGURE 10. M ) and is similar to that observed in specimens studied by Hayward (1988) and Thornely (1905). The shape of the leaf-like mandibles is also very constant among Red Sea and Indian Ocean specimens ( Figs 8 View FIGURE 8. M C, 11) and similar to that of mandibles illustrated by Harmer (1957, pl. 62, fig. 30, Kei Isl., Indonesia), Hayward (1988, pl. 10e, Mauritius) and Scholz et al. (2001, fig. 5c, Socotra). The only differences are observed among Lebanese colonies, which have mandibles with narrower wings and smaller frontal and marginal pores. In all our specimens, the lower face of the mandible presents pointed lateral processes on both sides of the proximal end of the rachis, which inserts in the channelled rostrum. However, when the broadest side of the mandible is particularly developed with a proximal lobe, the process corresponding to it on the back side is generally much less developed ( South Africa) or even missing ( Oman). Examination of numerous dry mandibles in the Oman and South African material showed that there is only one mandible type, but curling of their wings when drying often results in a narrow ‘setiform’ shape. This result contrasts with the dimorphism observed in colonies of M. harmeri by Shirakawa (1999, fide Taylor & Mawatari 2005). The shape and development of the ascopore are relatively variable within colonies and between localities ( Fig. 9 View FIGURE 9. M ). The median process was particularly developed and broad in specimens from Safaga Bay and South Africa. The presence in specimens from Bangladesh of orifices with a denticulate proximal rim, 3-4 oral spines and unpaired avicularia, among other characters, allowed Gordon et al. (2007) to assign them to M. harmeri . However, this attribution may be questionable in the absence of data on the mandible type and because the avicularian opesia appears to be smaller than in M. harmeri , such as in M. ventricosa Canu & Bassler, 1929 (see below).
Microporella harmeri also occurs in the Eastern Atlantic. This is evidenced by the precise illustrations and description of specimens from the Canary Islands attributed to M. orientalis by Aristegui (1984). These specimens have all typical features of M. harmeri including those associated with the ovicell (personate structure), the orifice (spines, denticulation) and the avicularium, particularly the broad opesia and the mandible with asymmetric lateral flaps and pointed processes on the lower face.
A specimen from the Gulf of Suez labelled as Microporella umbracula in the Balavoine Collection (MNHN n° 7803) is a Microporella species with personate maternal zooids and single avicularia with a broad lanceolate mandible. These features may correspond to M. harmeri .
The type of denticulate proximal edge and the shape of avicularium rostrum and mandible of specimens from Heron Island, Great Barrier Reef, attributed by Ryland and Hayward (1992) to M. orientalis suggested that they belong to M. harmeri . This was subsequently confirmed by checking unpublished SEM photos (courtesy of P.D. Taylor) of these specimens kept in the Natural History Museum, London (n° 1996.2.23.92).
Zabala and Maluquer (1988, p. 140, fig. 338) included in their key to the Mediterranean species of Microporella one that they ascribed to M. orientalis and characterized by personate ovicells (“expansions of the ovicell surrounding the aperture, often forming a tube rising above the orifice or curving towards the ascopore”) and single avicularia having “mandible setiform, with a pair of hook-shaped processes, situated halfway along its length”. Curiously, although their description was copied from Hayward (1974, p. 381), who gave no illustration of a single specimen from Chios ascribed to M. orientalis, Zabala and Maluquer illustrated their key with a drawing showing orifices with denticulate proximal edge, avicularia with particularly wide opesia and, in contradiction with their description, a mandible bearing two almost symmetrical lateral flaps. This Mediterranean (?) species, noted by Zabala and Maluquer (1988) as rare but without indication of origin, differs from that described by Hayward (1974); it is not M. orientalis and could belong to the M. harmeri species group.
Microporella harmeri View in CoL has many features in common with Microporelloides (Patorporella) hawaiiensis Soule, Chaney & Morris, 2003 View in CoL , particularly three spines, marginal areolae, denticulate proximal edge of orifice, personate maternal zooids with porous ooecia, and similarly shaped ascopore and avicularia. However, lanceolate mandibles were not noted in M. hawaiiensis View in CoL by Soule et al. (2003) and the ascopore is clearly distant from the personate collar instead of being adjacent to it as in M. harmeri View in CoL . Another similar species is Microporelloides (Microporelloides) lepueana Soule, Chaney & Morris, 2004 View in CoL , but it sometimes has paired frontal avicularia. These two Pacific species obviously constitute a species group with M. harmeri View in CoL . Considering the geographical distribution of the latter (Atlantic Ocean, Red Sea, Indian Ocean), it might be possible that several cryptic species remain to be identified.
Assessment of Microporella View in CoL specimens from the Albatross expedition to the Philippine Archipelago (Canu & Bassler 1929), in the USNM, Washington, revealed unexpected taxonomic connections. SEM examination revealed that several specimens ascribed to M. ciliata View in CoL (Canu & Bassler 1929, p. 331, pl. 40, figs 2–4) are conspecific with M. ventricosa Canu & Bassler, 1929 (p. 333, pl. 40, fig. 6), whose characters indicate evident similarities with M. harmeri View in CoL . The holotype of M. ventricosa (USNM 8109, Albatross Stn 5217) and specimens identified as M. ciliata View in CoL by Canu and Bassler (USNM 8103, 8104, 8105, 8106) display primary orifices with a denticulate proximal edge as in M. harmeri View in CoL , a feature not mentioned by Canu and Bassler, and other characters also observed in M. harmeri View in CoL (e.g. single avicularium, granular frontal wall). However, this set of specimens including M. ventricosa differs from M. harmeri View in CoL in having 4–5 strong, not ephemeral oral spines, ascopores surrounded by a cup-like prominent rim and avicularia with a smaller opesia. The average ratio of the width of the avicularian crossbar to the width of the orifice proximal edge calculated on SEM photos is about 0.6 in those Philippine specimens while it reaches 0.8–1 in M. harmeri View in CoL . The ancestrula seen in specimen 8105 has a more developed cryptocyst than in our specimens of M. harmeri View in CoL . Therefore, we consider that M. ventricosa differs significantly from M. harmeri View in CoL , but belongs with the latter and with M. hawaiiensis View in CoL and M. lepueana View in CoL in a distinct species complex. It also should be noted that the set of USNM specimens mentioned might belong to more than one species. Personate ovicells seen in 8103 have oral spines in the corners of the personate structure, whereas they are absent in the ovicells of 8106. There is a conspicuous rim around the avicularia in specimens 8103, 8104 and 8105, but not in 8106.
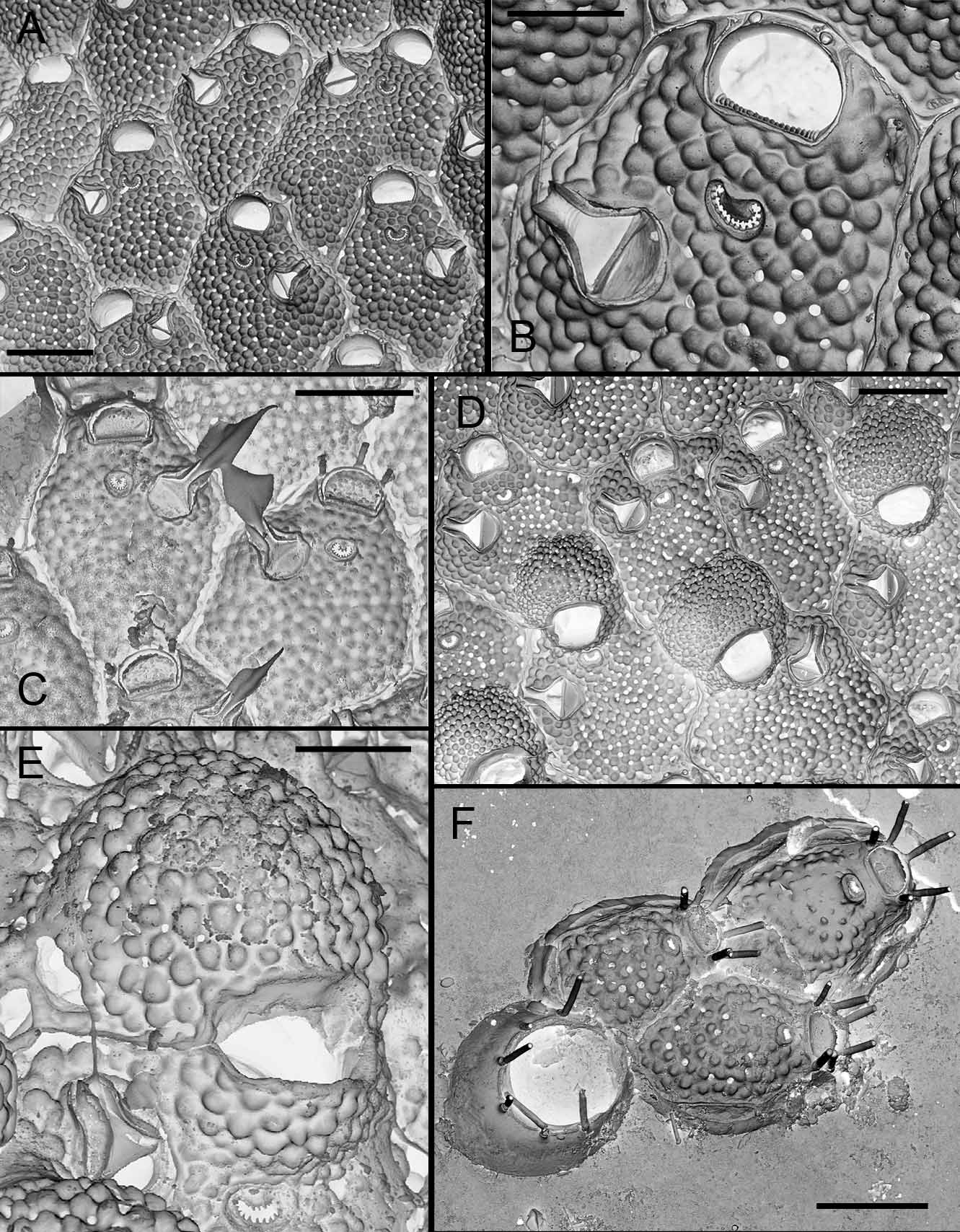
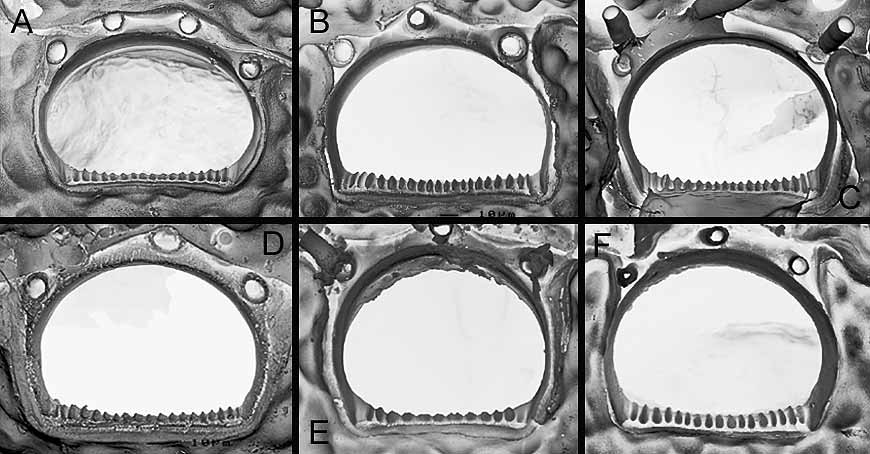
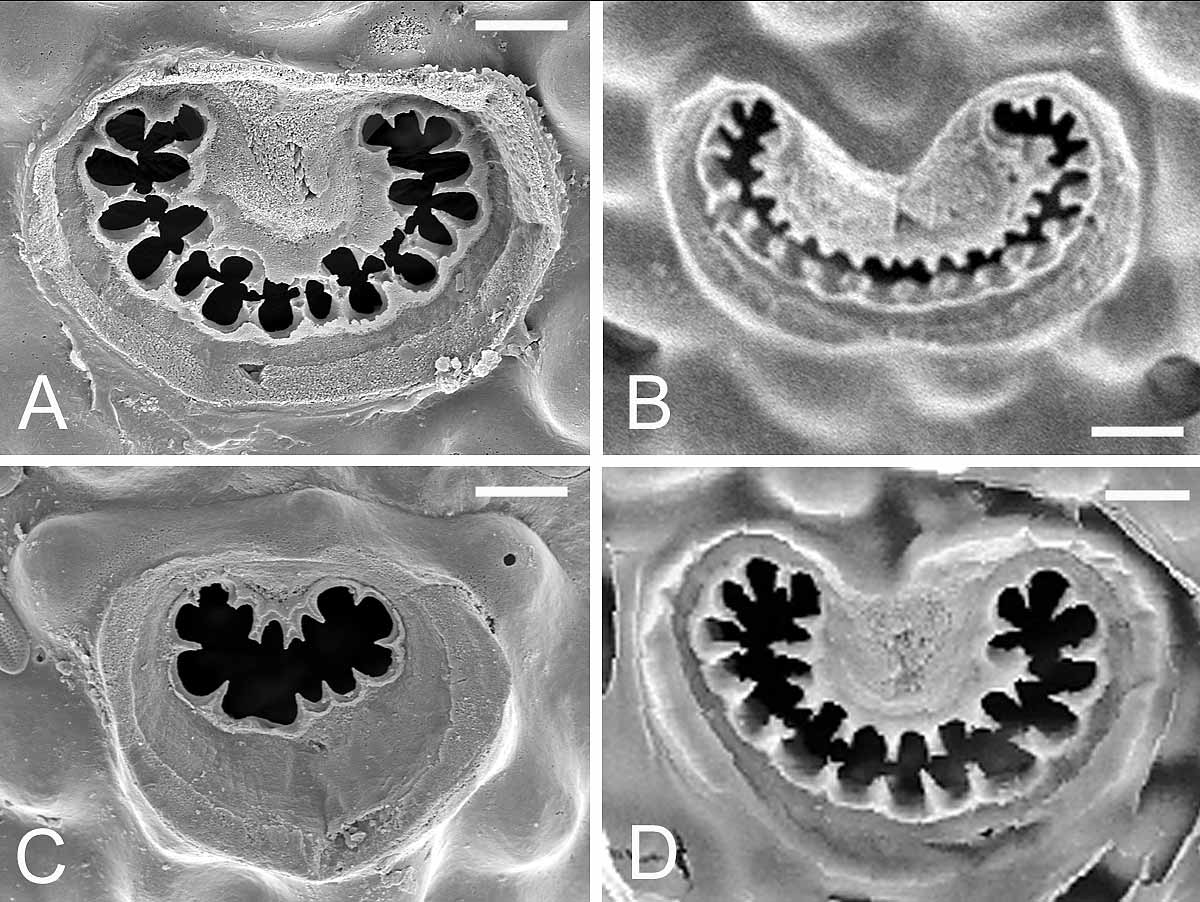
TABLE 5. Morphometrics (in µm) of specimens of M. harmeri from SE Mediterranean, Red Sea and Indian Ocean. Length (L) and width (W) of autozooid (Az), ovicell (Ov), primary orifice (Or) and avicularium mandible (Md). Mean standard deviation, range and number of measurements (in brackets).
| AzL 479.4 ±35.6 | 594.5 ±44.9 | 509.1± 44.7 | 562.3 ±34.8 | 687.2 ±59.7 | 522± 43.6 |
|---|---|---|---|---|---|
| - 440–540 (16) AzW 401.8± 55.5 | 535–660 (10) 506.5 ± 66.1 | 410–620 (65) 406± 49.5 | 470–630 (26) 417.3± 36.0 | 600–800 (18) 496.1± 46.0 | 440–600 /(20) 409.5 ±48.2 |
| - 300–485 (16) OvL 255.1 ± 25.0 - 205–300 (12) OvW 291.5 ± 20.4 - 240–320 (12) OrL 74.7 ± 3.3 | 435–630 (10) 292.5 ± 31.4 245–340 (10) 345.0 ± 19.7 315–365 (10) - | 310–550 (65) - - - - 80.6 ± 6.5 | 350–480 (26) 228.4 ± 19.2 180–260 (19) 278.9 ± 38.3 220–320 (19) 88.3 ± 4.0 | 410–580 (18) 179.4 ± 14.8 160–200 (16) 224.4 ± 8.9 210–240 (16) 84.4 ± 6.8 | 290–500 (20) - - - - 89.5 ± 4.8 |
| - 70–80 (6) OrW 109.4 ± 7.0 - 100–115 (9) MdL 188.8 ± 25.6 - 145–240 (17) | - 123.4 ± 4.7 117–128 (5) 273.1 ± 30.0 216–316 (9) | 72–90 (55) 106.7 ± 7.6 90–120 (55) 245 ± 16 210–260 (8) | 80–95 (26) 107.9 ± 7.1 90–120 (26) 258.7 ± 17.2 240–300(26) | 70–90 (18) 110.3 ± 8.1 100–120 (18) 260.3 ± 16.6 230–290 (15) | 80–100 (20) 120 ± 6.5 110–135 (20) 266 ± 30.6 210–300 (10) |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Microporella harmeri Hayward, 1988
| Harmelin, Jean-Georges, Ostrovsky, Andrew N., Cáceres-Chamizo, Julia P. & Sanner, Joann 2011 |
Microporelloides (Microporelloides) lepueana
| Soule, Chaney & Morris 2004 |
Microporelloides (Patorporella) hawaiiensis
| Soule, Chaney & Morris 2003 |
Microporella harmeri
| Hayward 1988 |
Microporella orientalis:
| Aristegui 1984 |
M. ventricosa
| Canu & Bassler 1929 |
var. personata:
| Thornely 1905 |