
KENYAPOTAMINAE AND EARLIEST
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00548.x |
|
persistent identifier |
https://treatment.plazi.org/id/03808792-FFB7-FF9B-FCBC-08C090F0FCE7 |
|
treatment provided by |
Valdenar |
|
scientific name |
KENYAPOTAMINAE AND EARLIEST |
| status |
|
KENYAPOTAMINAE AND EARLIEST HIPPOPOTAMINAE : AN AFRICAN HISTORY
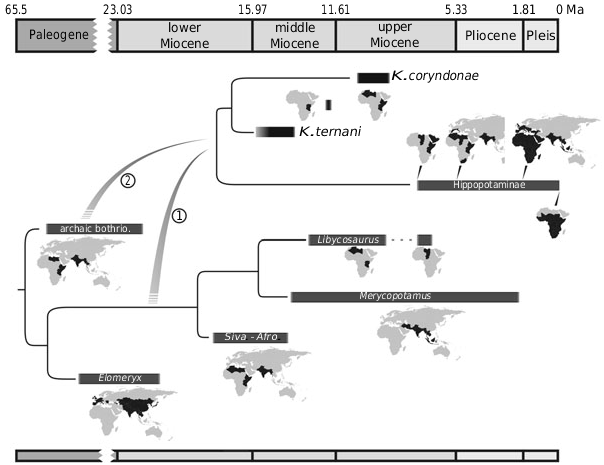
The known first 6 000 000 years of hippopotamid evolution was restricted to a limited portion of eastern Africa ( Fig. 11 View Figure 11 ). It is tempting to find some explanations for this situation beyond fossil record perfect- ibility. Hippopotaminae are characterized by a semiaquatic way of life constraining their dispersal abilities, arid areas being insurmountable barriers for them ( Jablonski, 2003). If Kenyapotaminae displayed a similar ecology, the phase of aridification experienced by Africa after 15 Mya ( Zachos et al., 2001) could be viewed as a factor limiting the distribution of the earliest hippopotamids (see also Boisserie & Lihoreau, 2006). Similarly, the later extension of kenyapotamines to central northern Africa ( Fig. 11 View Figure 11 ) should indicate the continuity of wet habitats within a ‘Trans-Saharan’ biogeographical province around 10 Mya. This scenario is supported by other data, notably on the semiaquatic Libycosaurus ( Lihoreau et al., 2006) .
Unfortunately, the ecology of kenyapotamines remains largely unknown. To date, the most reliable data come from analyses of enamel stable isotopic content performed by Harris et al. (2008) on some late Miocene kenyapotamine specimens. They notably show that the 18 O/ 16 O ratio, a parameter complexly linked to water dynamics in habitats and diets, is not significantly different in kenyapotamines and hippopotamines. This suggests similar semiaquatic habits in both subfamilies, but it is not possible to conclude firmly without comparing these stable isotope results with more general ones obtained for the complete faunas including Kenyapotamus .
The last known remains of Kenyapotamus predate the ‘hippopotamine event’. This event corresponds to the sudden appearance around 7.5 Mya of particularly abundant, fully evolved hippopotamines dominating local wet ecosystems in the Arabo-eastern African and northern central African fossil records (Boisserie, 2006; Weston & Boisserie, in press). This event may be an artefact of incomplete fossil records, the result of a punctuated equilibrium, a dramatic increase of hippopotamine abundance in relation to environmental changes, or a combination of these effects. Earliest known hippopotamines represented a considerable biomass in depositional environments, were relatively diversified, and were characterized by a distribution extended at least to the northern half of the continent. These elements seem in favour of an emergence of ‘true’ hippopotamines well before the ‘hippopotamine event’. Morphology tends to support this view: late Miocene kenyapotamines seem indeed a less suitable stem group for Hippopotaminae than middle Miocene ones, suggesting a deeper hippopotamine evolutionary history. Resolution of this question will probably come from new discoveries, either of a form transitional between K. coryndonae and late Miocene hippopotamines, or, more likely in our opinion, of a hippopotamine lineage contemporary to the late Miocene Kenyapotamus .
Expansion of Hippopotamidae outside Africa is recorded only at the terminal Miocene, after the ‘hippopotamine event’, to southern Europe and southern Asia ( Fig. 11 View Figure 11 ). Maximal distribution of the family was reached during the Pleistocene, with Pan-African distribution, presence of Hippopotamus in western to south-eastern European and south-western Asia, and extension of Hexaprotodon from southern Asia to south-eastern Asia ( Kahlke, 1990; Boisserie, 2007). It would be important to look for Mio-Pliocene hippopotamid remains in south-western Asia in order to complete the biogeographical history of the family.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |