
Thescelosauridae Sternberg, 1937
|
publication ID |
https://doi.org/ 10.26879/702 |
|
persistent identifier |
https://treatment.plazi.org/id/6D2D8786-A268-5F0C-741E-FD4F26804DBE |
|
treatment provided by |
Felipe |
|
scientific name |
Thescelosauridae Sternberg, 1937 |
| status |
|
Thescelosauridae Sternberg, 1937
Phylogenetic definition. All neornithischians more closely related to Thescelosaurus neglectus Gilmore, 1913 than to Hypsilophodon foxii Huxley, 1869 , Dryosaurus altus (Marsh, 1878) , or Parasaurolophus walkeri Parks, 1922 .
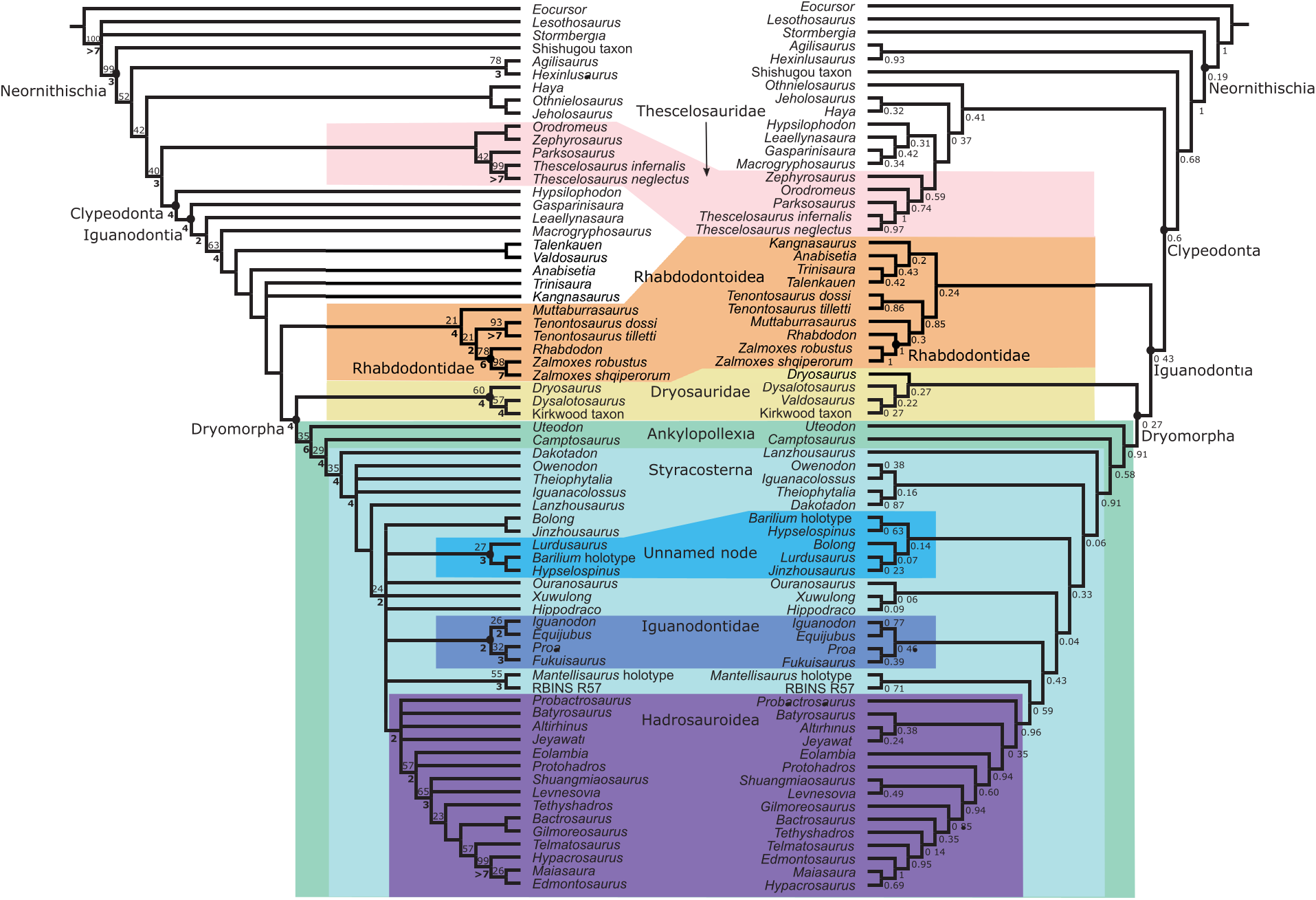
Unambiguous synapomorphies. In the parsimony analysis ( Figure 7 View FIGURE 7 ), thescelosaurids are characterized by three unambiguous synapomorphies: dentary tooth row straight in dorsal view (98.0), 10-12 ridges on the maxillary teeth (136.3), and acromion process of scapula does not extend beyond the edge of the coracoid (196.0).
In the Bayesian analysis ( Figure 8 View FIGURE 8 ), a more extensive Thescelosauridae is characterized by two unambiguous synapomorphies: 10-12 ridges on the maxillary teeth (136.3) and 9-11 ridges on the dentary teeth (137.3).
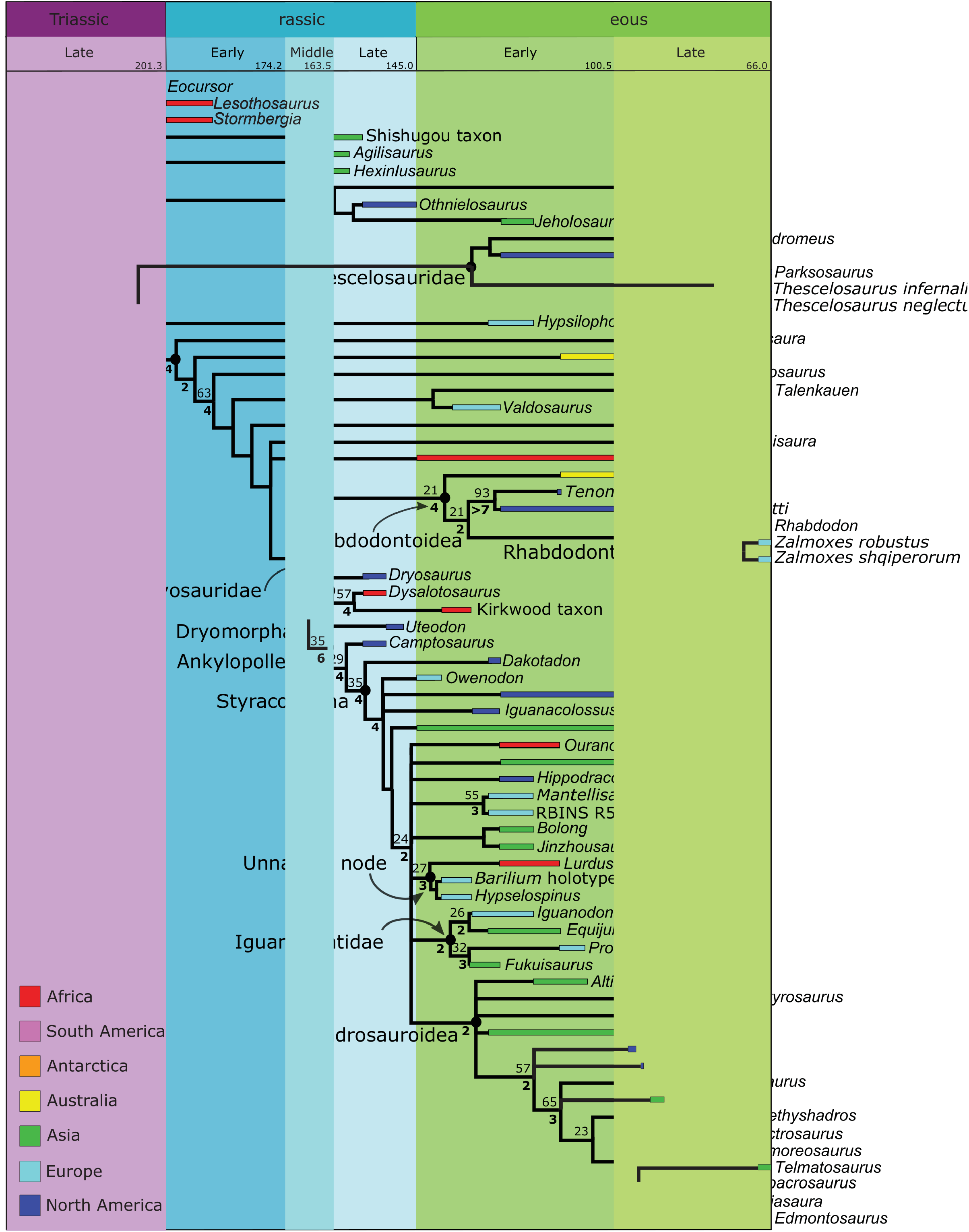
Topology. Thescelosauridae is recovered in the parsimony analysis ( Figures 6 View FIGURE 6 , 7 View FIGURE 7 ) containing Thescelosaurus , Parksosaurus , Orodromeus , and Zephyrosaurus , though the group has poor support (Jackknife value=7). Unlike in Boyd (2012), Haya is recovered at a node basal to thescelosaurids, rather than within the Thescelosaurinae. Macrogryphosaurus and Talenkauen , which Boyd also found within Thescelosaurinae, are recovered within iguanodontia, based on the presence of opisthocoelus cervical vertebrae, and a postacetabular process that is less than 30% of total ilium length.
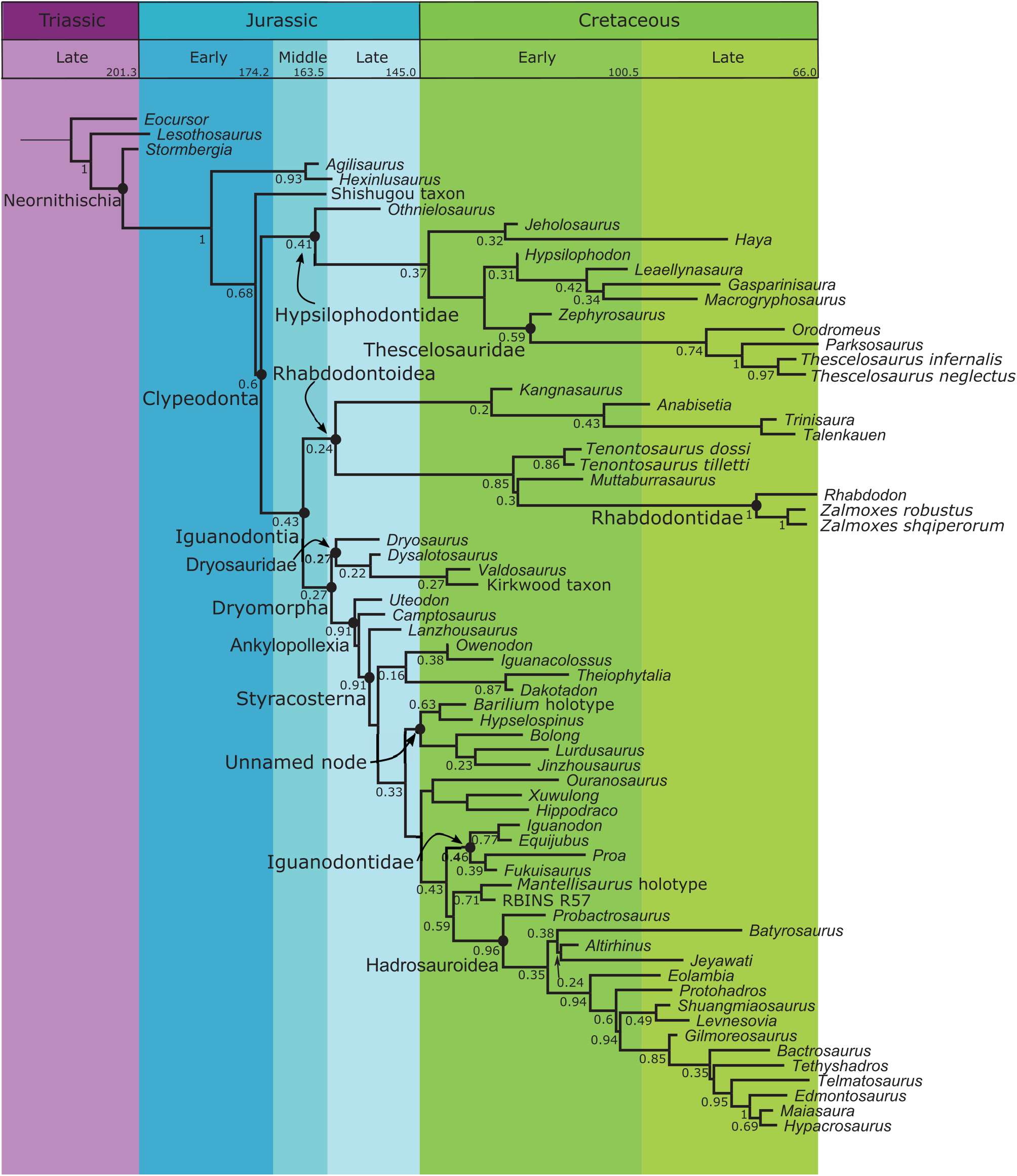
In the Bayesian analysis ( Figures 6 View FIGURE 6 , 8 View FIGURE 8 ), Thescelosauridae is nested within a larger Hypsilophodontidae . Though it only has moderate support (PP=0.59), the internal nodes have higher posterior probabilities. The genera included in this group match those recovered in the parsimony analysis; the one difference in topology is that Orodromeus and Zephyrosaurus form successive outgroups to Thescelosaurus + Parksosaurus instead of forming a distinct Orodrominae (as found by Boyd 2012). Though Haya and Macrogryphosaurus are recovered as closely related hypsilophodontids, they are not found within Thescelosauridae as proposed in Boyd (2012).
CLYPEODONTA Norman, 2015
Phylogenetic definition. Hypsilophodon foxii , Edmontosaurus regalis , their most recent common ancestor, and all of its descendants (Norman, 2015).
Unambiguous synapomorphies. For the topology recovered by parsimony, Clypeodonta has nine unambiguous synapomorphies: presence of a quadrate buttress or "hamular process" (63.1), quadrate with a lateral condyle that is larger than the medial condyle (69.2), mandibular articulation that is horizontal to dorsomedially inclined in caudal view (70.0/1), maxillary and dentary teeth with crowns that taper toward the root (127.1, 128.1), with the base of the crown defined by an everted lip which makes the crown slightly inset from the root (146.1, 147.1), presence of a primary ridge on labial side of maxillary teeth (139.1), and elongate centra of postaxial cervical vertebrae, with craniocaudal length more than twice the dorsoventral height (159.1).
Within the Bayesian topology, Clypeodonta is characterized primarily by features of the teeth and jaws: presence of a diastema in the maxilla (16.1), equal lengths in the oral margin of the premaxilla and predentary (84.1), a coronoid process that extends more than one crown height dorsal to the tooth row (101.1), surangular with a small fenestra positioned dorsally on or near the dentary joint (111.1), surangular foramen rostral to the lateral lip of the glenoid (114.1), cheek teeth with asymmetrically distributed enamel (134.1), ridges running the full length of the crown on the labial side of maxillary teeth and the lingual side of dentary teeth (135.1), and a femoral head separated from the greater trochanter by a distinct constriction (292.1). Topology. In the parsimony tree ( Figures 6 View FIGURE 6 , 7 View FIGURE 7 ), Clypeodonta is the sister clade to Thescelosauridae . This node has a jackknife value of 15, but a relatively high Bremer support of 4. Hypsilophodon is recovered as the only non-iguanodontian clypeodontan.
In the Bayesian topology ( Figures 6 View FIGURE 6 , 8 View FIGURE 8 ), Hypsilophodon is recovered within a large Hypsilophodontidae . Consequently, Clypeodonta is a more inclusive clade than in the parsimony tree, including Thescelosauridae , a clade with Haya , Jeholosaurus , and Othnielosaurus , and Leaellynasaura , Gasparinisaura , and Macrogryphosaurus . It excludes only a few basal neornithischians such as Hexinlusaurus and Agilisaurus . It is moderately supported, with a posterior probability (PP) of 0.60.
IGUANODONTIA Sereno, 1986
Phylogenetic definition. The most inclusive clade containing Parasaurolophus walkeri Parks, 1922 but not Hypsilophodon foxii Huxley, 1869 , or Thescelosaurus neglectus Gilmore, 1913 (Sereno, 2005).
Unambiguous synapomorphies. In the parsimony analysis, Iguanodontia is characterized by a maxilla with a broad and triangular dorsal process of the maxilla (21.1), a quadrate extending ventrally such that the quadratojugal is well removed from the mandibular condyle (60.1), a single wear facet on each cheek tooth (131.1), opisthocoelus post-axial cervical vertebrae (157.1), a distinct indentation on the scapula superior to the glenoid, termed here the supraglenoid fossa (199.1), and a manual digit III with three or fewer phalanges (236.1).
Two other synapomorphies recovered for this clade are elongate prezygopophyses on the distal caudal vertebrae (185.1), and chevrons that are strongly and asymmetrically expanded distally (188.1). The former of these is found only in Gasparinisaura and Leaellynasaura , and the latter in these genera plus Parksosaurus and Macrogryphosaurus . While they are present at the base of the clade, these characters are not widespread, and therefore not useful in diagnosing the clade.
There is only one overlapping character here with the diagnosis of Sereno (1986); the reduction of phalanges in digit III. The presence of “leaf-shaped” or mamillated denticles is more restricted within Styracosterna, and while most iguanodontians lack premaxillary teeth, both Talenkauen and Tenontosaurus dossi have one premaxillary tooth. Iguanodontia is recovered with jackknife support of 19 and Bremer support of 4.
Within the Bayesian topology, Iguanodontia (PP=0.43) lacks the basal pectinate region found in the parsimony analysis and is instead composed of the sister groups of rhabdodontoids and Dryomorpha. Gasparinisaura , Leaellynasaura , and Macrogryphosaurus are excluded from Iguanodontia, and are recovered instead with the hypsilophodontids. This rearrangement of taxa leads to different synapomorphies for Iguanodontia between the parsimony and Bayesian analyses. Synapomorphies for the Bayesian topology include: premaxilla flaring laterally to form a floor of the narial fossa (3.1), small antorbital fenestra (31.1), predentary with denticulate oral margin (87.1), ventral process of predentary deeply bifurcated (89.1), cheek teeth with crowns tapering toward the root (127.1, 128.1), cheek teeth that are closely packed without spaces between roots (126.1), cheek teeth with one wear facet on each tooth (131.1), cheek teeth lacking a basal ridge (“cingulum”) (148.1), caudal vertebrae with distal facets for chevrons much larger than proximal facets (183.0), humerus with an elongate deltopectoral crest (>43% humeral length) (214.1), manual digit III with three or fewer phalanges (236.1), first phalanx of manual digits II-IV more than twice the length of the second phalanx (239.1), ischium with an untwisted shaft (283.1), ischium with an expanded distal end (288.1), femur with a cranial intercondylar sulcus (300.1), and a caudal intercondylar sulcus partially roofed by the medial condyle (302.1).
Topology. In the parsimony analysis ( Figures 6 View FIGURE 6 , 7 View FIGURE 7 ), the basally branching portion of Iguanodontia forms a pectinate topology outside of Dryomorpha, which includes Gasparinisaura , Leaellynasaura , Macrogryphosaurus , Talenkauen , Valdosaurus , Anabisetia , Trinisaura , and Kangnasaurus . It is supported by a jackknife value of 19 and a Bremmer support of 4.
In the Bayesian analysis ( Figures 6 View FIGURE 6 , 8 View FIGURE 8 ), Iguanodontia bifurcates into rhabdodontoids and dryomorphans. Iguanodontia is supported by a posterior probability of 0.43.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.