
Dendrocoelum leporii Stocchino & Sluys, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4323.4.7 |
|
publication LSID |
lsid:zoobank.org:pub:69Fa8637-8007-4216-80Ee-8A261B827Abf |
|
DOI |
https://doi.org/10.5281/zenodo.6021639 |
|
persistent identifier |
https://treatment.plazi.org/id/03848789-EA7F-E242-FF78-FAD0FE3F4183 |
|
treatment provided by |
Plazi |
|
scientific name |
Dendrocoelum leporii Stocchino & Sluys |
| status |
sp. nov. |
Dendrocoelum leporii Stocchino & Sluys , sp. nov.
Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ; Table 1
Diagnosis. Dendrocoelum leporii is characterized by: a muscular atrial valve; a penis papilla with a pseudoflagellum; numerous testes situated in the ventral, dorsal and median body regions and extending to the far posterior end of the body; a large adenodactyl with the so-called Balkan type of musculature.
Etymology. The new species is dedicated to the late Prof. Dr. Nullo Glauco Lepori who was eminent in the field of biological and systematic studies on freshwater triclads.
Material examined. Holotype: ZMA V.Pl. 7271.1, Sorgente del Tunnel, Frasassi cave complex, Frasassi Gorge (43°24’04.53” N, 12°57’57.14” E), Italy, 19 March 2011, coll. A. Montanari, sagittal sections on 6 slides. GoogleMaps
Other material: CGAS Pla 17. 1, Sorgente del Tunnel , Frasassi cave complex, Frasassi Gorge (43°24’04.53” N, 12°57’57.14” E), Italy, January 2014, coll. A. Montanari, sagittal sections on 9 slides. GoogleMaps
ZMA V.Pl. 7272.1, Pozzo dei Cristalli, Grotta del Fiume, Frasassi cave complex (43°24’02.27” N, 12°57’53.17” E), Italy, 19 March 2012, coll. A. Montanari, sagittal sections on 6 slides. GoogleMaps
Habitat. The Frasassi cave complex encompasses some 100 caves of hypogenic origin, resulting from the interaction between limestone and sulfidic waters ascending from a deep aquifer (cf. Sarbu et al., 2000) ( Fig. 2 View FIGURE 2 ). The cave system forms a ramifying pattern at different, overlying levels (cf. Sarbu et al., 2000; Mariani et al., 2007 and references therein) ( Fig. 2B View FIGURE 2 ). The water of the streams and lakes in this cave system is characterized by greatly different concentrations of H2S and O2 (cf. Macalady et al., 2008; Flot et al., 2010). The Frasassi cave complex houses the most visited show cave of Italy, viz. the Grotta Grande del Vento ( Fig. 2A View FIGURE 2 ).
Stygofauna reported for the Frasassi cave complex includes Nematoda, Annelida ( Naididae ), Rotifera (Bdelloidea), Mollusca (Gastropoda), Crustacea (Ostracoda, Copepoda, and Amphipoda ), and Platyhelminthes (Tricladida and Catenulida) ( Bertolani et al., 1994; Dattagupta et al., 2009; Karaman et al., 2010; Montanari, 2010; Bodon & Cianfanelli, 2012; Peterson et al., 2012; Bauermeister et al., 2013; Galassi et al., 2017).
The planarians described in this paper were found in the subterranean spring Sorgente del Tunnel and in the Pozzo dei Cristalli section of the Grotta del Fiume cave, both being parts of the Frasassi cave complex. However, several other specimens of dendrocoelids were observed at other sites of the Frasassi cave complex, namely the Grotta Sulfurea, the Ramo Solfureo section in the Grotta del Fiume (S. Dattagupta pers. comm.), and at Lago Claudia in the innermost part of the Grotta Grande del Vento (D. Peterson pers. comm.) ( Fig. 2A View FIGURE 2 ).
Sorgente del Tunnel is a sulfidic spring flowing inside a man-made tunnel, located on the hydrographic right bank of the Sentino River near the eastern end of the Frasassi Gorge ( Fig. 2A View FIGURE 2 ). Planarians were found moving on the black sulfidic mud of the very shallow rivulet of the spring at about 20 m from the entrance of the tunnel. Water depth: 2-5 cm; water temperature: ca. 13.5 °C all year round; pH: ca. 7. Unfortunately, at present the spring and the tunnel are inaccessible as a result of a landslide that obstructs the entrance since the Sentino River was heavily flooded in 2013.
The entrance of Grotta del Fiume is located on the right bank of the Sentino River in the Frasassi Gorge, ca. 3 m above river level (at 208 m asl) ( Fig. 2A View FIGURE 2 ). Planarians were found on the black sulfidic mud of the Pozzo dei Cristalli section of the cave ( Fig. 2A View FIGURE 2 ), mixed with filaments of sulphur-oxidizing bacteria, in a small, shallow pool close to a flowing stream of sulfidic water. Water depth: 2-5 cm; water temperature: ca. 13.5 °C; pH: 7; H2S: 30 ppm.
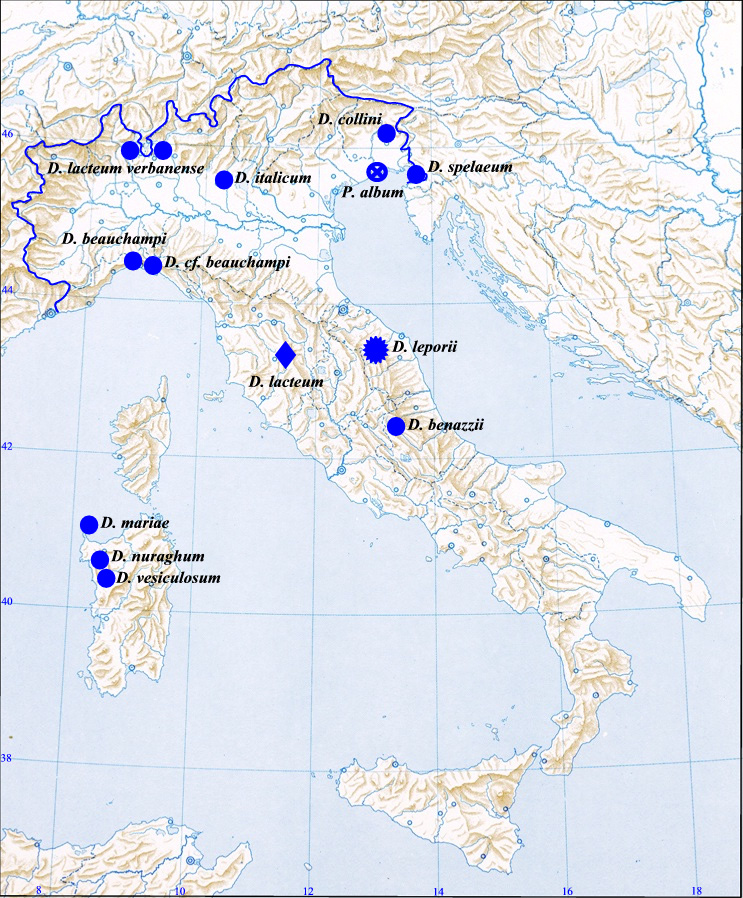
Geographical distribution. Exclusively known from the Frasassi cave complex in central Italy.
Description. Live animals unpigmented, typically whitish, and lacking eyes. Live mature specimens with a body length of 10–15 mm and a width ranging from ca. 3 mm in the central part of the body to ca. 1 mm at the level of the head. Anterior end truncated, with the middle part of the frontal margin being convex and provided with a pair of small, rounded lateral lobes ( Figs 3 View FIGURE 3 , 4 View FIGURE 4 ).
Subterminal anterior adhesive organ moderately developed, consisting of a shallow cup made up of infranucleated epithelial cells, which are pierced by numerous openings of erythrophil glands. The musculature associated with this organ consists of a more strongly developed part of the usual ventral longitudinal body musculature.
The pharynx is located in the middle of the body and measures about 1/8th of the body length ( Figs 3 View FIGURE 3 , 4 View FIGURE 4 ). Its internal muscle zone consists of a very thick layer of intermingled circular and longitudinal fibres. A subepithelial layer of longitudinal muscles, followed by a layer of circular fibres, forms the thin outer zone of pharyngeal musculature.
The ventral ovaries are located at 1/4th of the distance between the brain and the root of the pharynx. The oviducts originate from the antero-lateral part of the ovaries and are provided with an expansion at their anterior end, thus forming a tuba. The oviducts, which are lined with an infranucleated epithelium, run posteriorly, recurve posterior to the gonopore and, subsequently, fuse to form a common oviduct. The latter runs anteriad to the right side of the bursal canal to open into the distal, ventro-posterior part of the male atrium. The common oviduct receives the openings of erythrophil shell glands along the posterior half of its length ( Fig. 5A View FIGURE 5 ).
Well-developed, rounded resorptive vesicles are present along the oviducts of all specimens examined, while they are particularly numerous in specimen ZMA V.Pl. 7272.1. Each vesicle communicates with the oviduct through a short, narrow ductule. The latter is localized very close to the ductule connecting each vitellarium with the oviduct ( Fig. 6C View FIGURE 6 ).
Well-developed, numerous testes are present in both dorsal and ventral position, while some follicles occupy the entire dorso-ventral diameter of the body. The testes extend from a short distance behind the ovaries far into the posterior end of the body.
In specimens ZMA V.Pl. 7272.1 and CGAS Pla 17. 1 the sperm ducts form well-developed spermiducal vesicles, located between the mouth and the anterior level of the penis bulb, which are packed with sperm. The sperm ducts curve towards the dorsal body surface and, subsequently, separately penetrate the antero-lateral surface of the penis bulb to open symmetrically and very closely together into the anterior section of the seminal vesicle ( Fig. 5A View FIGURE 5 ).
The copulatory apparatus occupies the anterior half of the postpharyngeal region. The sac-shaped copulatory bursa occupies the entire dorso-ventral space of the body just behind the pharyngeal pouch. The bursa is lined with a cuboidal to columnar glandular epithelium and is surrounded by a sheath of muscles ( Fig. 5A–B View FIGURE 5 ).
From its point of communication with the bursa the bursal canal runs posteriorly and horizontally to the left of the penis. This narrow horizontal tract of the bursal canal is lined with a nucleated, ciliated epithelium and surrounded by only a thin layer of longitudinal muscle fibres. At the level of the insertion of the adenodactyl papilla the bursal canal widens for a short tract and then it reduces again in diameter. From this last point onwards the lumen of the bursal canal increases again in diameter, while it is surrounded by a few layers of circular muscle, followed by a single layer of longitudinal fibres ( Fig. 5A–B View FIGURE 5 ).
The penis is located dorsally to the right of the adenodactyl ( Fig. 5A–B View FIGURE 5 ). In the holotype the penis is approximately 2/3rd of the length of the adenodactyl, while in specimens CGAS Pla 17. 1 and ZMA V.Pl. 7272.1 the penis has the same length as the adenodactyl. This variability in penial length depends on the state of contraction of both the penis and the adenodactyl.
The muscular penis bulb is well-developed and houses a narrow seminal vesicle. The latter communicates with the ejaculatory duct, which opens at the tip of the penis papilla. The dorsally displaced ejaculatory duct is lined with a nucleated epithelium and surrounded by very thin layer of longitudinal muscle fibres.
The basal part of the penis papilla is covered with a thin epithelium, which is underlain by a thick layer of circular muscle, followed by a layer of longitudinal fibres.
At the distal part of the penis papilla the epithelium covering the papilla becomes thicker, while the underlying musculature completely disappears.
Penis glands open into the terminal tract of the ejaculatory duct and at the apical portion of the penis papilla. The ejaculatory duct is full of secretion, as is the male atrium ( Fig. 6A–B View FIGURE 6 ).
In the holotype and in specimen CGAS Pla 17. 1 the penis papilla is conical, whereas in ZMA V.Pl. 7272.1 its shape is more elongated, depending on the state of contraction ( Figs 5A View FIGURE 5 , 6A–B View FIGURE 6 ).
The adenodactyl is very large and consists of a free papilla and a well-developed bulbar part. In the holotype and in specimen ZMA V.Pl. 7272.1 the adenodactyl is thrust out of the body, a condition very likely due to preservation artefacts. The bulb consists of intermingled rows of longitudinal and circular muscle, bounded by a thin layer of longitudinal fibres. The lumen of the adenodactyl, lined by a layer of ciliated cells, is surrounded by a well-developed zone of mesenchymatic tissue. Through this section of the mesenchyme runs a thick layer of fine circular muscle fibres. Ectally and internally to this zone of circular muscles runs a thin layer of longitudinal muscle fibres ( Figs 5A–B View FIGURE 5 ); this anatomy corresponds to the so-called Balkan type of adenodactyl.
The male atrium, which opens into the common atrium, is lined by a columnar, nucleated epithelium and is surrounded by a subepithelial layer of circular muscles, followed by a layer of longitudinal fibres.
Very close to the base of the penis papilla the circular muscles thicken even further, thus forming a muscular annular atrial fold or valve, which is more developed in its ventral portion than in its dorsal section in all specimens examined ( Figs 5A View FIGURE 5 , 6A–B View FIGURE 6 ).





| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |