
Crematogaster undetermined
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlab047 |
|
publication LSID |
lsid:zoobank.org:pub:6144DD31-0F7B-4589-86A3-F40994452C9 |
|
persistent identifier |
https://treatment.plazi.org/id/039987E6-FFDF-FFF7-FC31-7E3FFC196349 |
|
treatment provided by |
Plazi |
|
scientific name |
Crematogaster undetermined |
| status |
|
DELIMITING CREMATOGASTER SPECIES
In investigating the taxonomy of Crematogaster , we adopt the viewpoint that for sexual organisms, such as these ants, the biological species concept ( Mayr, 1942) provides a useful framework for understanding the nature of species and their morphological cohesiveness ( Coyne & Orr, 2004; Barraclough, 2019). At the same time, widely distributed species with large effective population sizes can be expected to harbour considerable phenotypic variation, distributed nonrandomly across the landscape. Delimiting species then involves the discovery of phenotypic or genetic gaps that indicate the occurrence of reproductive isolation among these population assemblages. This exercise requires comprehensive geographic sampling and a careful consideration of the observed variation. Morereover, because speciation is a continuous process, with complex dynamics at intermediate stages ( Noor & Bennett, 2009; Roux et al., 2016; Taylor & Larson, 2019) the findings may be properly ambiguous.
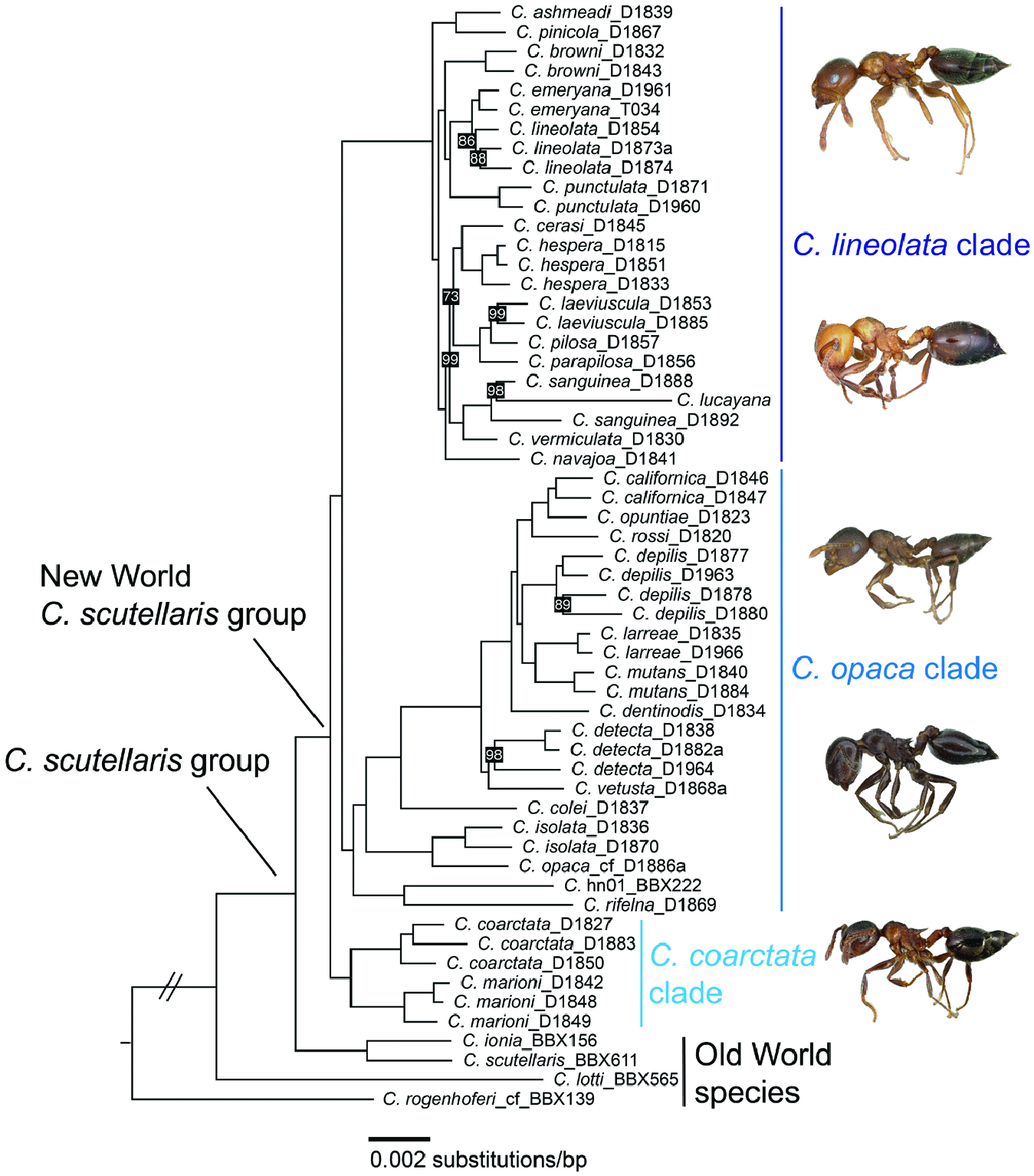
The results presented here, based primarily on morphology, should be considered tentative and subject to refinement with further study, especially more detailed genetic analyses. Whereas our morphological study of these Nearctic Crematogaster is based on examination of thousands of workers, the phylogenomic sample sizes are much smaller, in the range of one to four individuals per species. Nevertheless, we chose a geographically representative set of specimens for sequencing and there were instances in which the two sources of data were reciprocally illuminating. For example, our reinstatement – based on morphological evidence – of previously synonymized Crematogaster species was bolstered by the discovery that they did not have a close phylogenetic relationship to the species under which they had been synonymized. Conversely, where the molecular data indicated that two forms were sister-taxa – as we found for C. emeryana and C. lineolata – this directed attention to their geographical distributions, because maintenance of morphological differences in sympatry provides a more compelling case for treating two taxa as different species than if they are allopatric ( Galtier, 2019). In one instance, the phylogenomic data revealed more than one species among samples that we initially treated as conspecific: collections of C. detecta were originally misidentified as C. depilis , a species situated several nodes distant in the phylogeny ( Fig. 1 View Figure 1 ). Once their genetic distinctness was discovered, this motivated a search for morphological differences.
CONTINUING CHALLENGES IN CREMATOGASTER TAXONOMY
Because the C. scutellaris group is relatively young, with most diversification in the New World occurring within the last 5 Myr ( Fig. 2 View Figure 2 ), one can expect there to be ongoing differentiation of populations and incomplete reproductive isolation between some taxa, making clear delineation of species boundaries challenging. The preceding species accounts and the workerbased key to the North American representatives of the C. scutellaris group serve to summarize our current understanding of these ants, but there is still considerable taxonomic uncertainty about species limits.
The distinction between the allopatric taxa C. californica and C. opuntiae is a tenuous one, relying on a character (appressed scape pubescence in C. opuntiae ) that shows substantial variation. Crematogaster opuntiae may prove to be nothing more than a desert-dwelling variant of C. californica . Resolution of this issue requires further genetic and morphological studies, especially of populations from geographically intermediate areas and from farther south in Baja California. Another problematic taxon in this complex is C. nocturna, described from distinctivelooking males and alate queens from northern Arizona, but without associated workers. Buren (1968) identified a small series of six workers from southern Utah (not seen by us) as being possibly conspecific. Until the phylogenetic position of C. nocturna is determined, and queens and males are collected in association with workers, there will be ongoing uncertainty about the status of this species. There is also the challenging problem of distinguishing workers of C. depilis , C. detecta and C. larreae , discussed above but not fully resolved.
We let stand the synonymy by Morgan & Mackay (2017) of C. mormonum (type locality Utah) under C. coarctata (type locality San Francisco, California). This is one of the few synonymies in Morgan & Mackay (2017) that appears to be justified. The putative distinction in cephalic sculpture between these two forms – with shiny heads of C. mormonum workers being contrasted with the densely reticulate-foveolate heads of C. coarctata workers ( Buren, 1968) – is undermined by intermediate conditions among many individuals from northern California and Nevada. Nevertheless, among samples that we are now calling C. coarctata , there is considerable variation in shape, pilosity and sculpture. In particular, workers from southern California and northern Baja California tend to have reduced pronotal pilosity, a weakly sculptured promesonotum and shiny (C. mormonum -like) heads. Further study of this complex of populations is warranted.
Several investigators have commented on the extensive variation in colour and other features (pilosity, sculpture) that occurs within some species of Crematogaster from the eastern United States ( Creighton, 1950; Johnson, 1988; Deyrup, 2017). One such colour variant has been cleaved from C. ashmeadi and described as a distinct species, C.pinicola ( Deyrup & Cover, 2007), which differs in colour and nesting habits from the more widespread C. ashmeadi. The taxonomic significance of colour variation in other species, such as C. laeviuscula , C. parapilosa and C. pilosa , remains unclear. Among other problems affecting Crematogaster from eastern and central United States, the relationship of C. punctulata to C. lineoleata also requires further scrutiny, as discussed above.
The species-level taxonomy of the Caribbean representatives of Crematogaster (Crematogaster) is a further source of confusion.There are seven valid species or subspecies names (C. lucayana, C. lucayana etiolata Wheeler, 1905, C. manni Buren, 1968, C. sanguinea, C. sanguinea atavista Mann, 1920, C. sanguinea sotolongoi Mann, 1920 and C. sanguinea torrei Wheeler, 1913), but we have no clear understanding of the number of species or their relationship to one another. Our knowledge of the C. scutellaris group in Mexico and Central America is even more fragmentary. There are five valid names of taxa occurring almost entirely south of the United States in this region ( C. cedrosensis , C. opaca , C. patei Buren, 1968, C. rossi and C. saussurei Forel, 1899), but this belies a greater diversity that remains undersampled and understudied. The C. opaca complex, discussed above, is particularly troublesome and requires comprehensive morphological and phylogenetic investigation to clarify how many species exist and their boundaries.
Finally, there is a mysterious species, C. clydia Forel, 1912, known only from the holotype (unique syntype) worker collected more than one-hundred years ago in São Paulo state, Brazil. This is a Crematogaster (Crematogaster) species and presumably a member of the C. scutellaris group, and yet it represents an extraordinary out-of-range occurrence. Assuming that it has not been mislabelled, the rediscovery of C. clydia and clarification of its relationship to other congeners poses an intriguing challenge to Brazilian myrmecologists.
LOCATION OF TYPE SPECIMENS
In his treatment of the North American representatives of Crematogaster s.s. (i.e. the C. scutellaris group), Buren (1968) described 14 new species. He stated his intention to deposit the holotypes of the new species in the USNM, except those of C. nocturna and C. rossi , which were destined for CASC, and that of C. opuntiae , for which no place of deposition was indicated. However, Buren did not follow through for most species, so the information in his paper is misleading. Most holotypes were retained in his collection and that collection was acquired by the LACM in 1983, after Buren’s death. Through visits and loans we have determined the location of the holotypes of all 14 species and we provide this information here ( Table 3) for the benefit of future researchers. It will be noted that eight of the holotypes are in LACM, five in USNM and one in CASC.
Statements in Morgan & Mackay (2017) about the location of type specimens should also be viewed with caution. The authors use at least four collection acronyms (MNHG, NHMG, NMHG and NNHW) that are nowhere explained and are not part of the list of collections cited in the ‘Material and methods’ section. ‘MNHG’ appears to be a misspelling of MHNG (Muséum d’Histoire Naturelle, Genève), but some claims about types existing in that collection are incorrect, as documented above .
In fact, there are many errors and mistatements in the Morgan & Mackay (2017) monograph, particularly regarding the distribution and biology of particular species. The paper was published by a print-on-demand publisher, whose policy is to print without peer review or editorial supervision ( Fang, 2014; Stromberg, 2014; Anonymous, 2015). Taxonomy inevitably suffers if researchers pursue this kind of publication strategy, bypassing editor-mediated peer review and even the most perfunctory editing, and burdening other taxonomists with the task of correcting their mistakes.
NEW INSIGHTS INTO THE COMPOSITION AND ORIGIN OF THE CREMATOGASTER SCUTELLARIS GROUP
The Crematogaster scutellaris View in CoL group was circumscribed by Blaimer (2012b) as likely encompassing all Nearctic and Palaearctic representatives within the subgenus Crematogaster View in CoL , as well as a few species distributed in the northern Neotropical realm. This assumption was based on phylogenetic data from multiple nuclear loci ( Blaimer, 2012a) and morphological similarities of eight species (six Nearctic and two Palaearctic) within the C. scutellaris View in CoL group. That study indicated a clear phylogenetic division between a Nearctic and a Palaearctic clade ( Blaimer, 2012a). Our phylogenomic study has expanded taxon sampling within the C. scutellaris View in CoL group to include nearly all the Nearctic members of the group and a representative sampling of Caribbean and northern Neotropical taxa. Our results reveal a strongly supported New World clade, which is sister to the two sequenced Palaearctic species. A comprehensive overview of the C. scutellaris View in CoL group will only be possible once the phylogenetic affinities of the Mexican/Mesoamerican and remaining Palaearctic Crematogaster (Crematogaster) species are tested. In contrast to the tight relationships of the subgenus in the New World, it appears likely that some Palaearctic species are not members of the C. scutellaris View in CoL group (sensu Blaimer, 2012b), but more closely related to African Crematogaster View in CoL lineages (Blaimer, unpublished data).
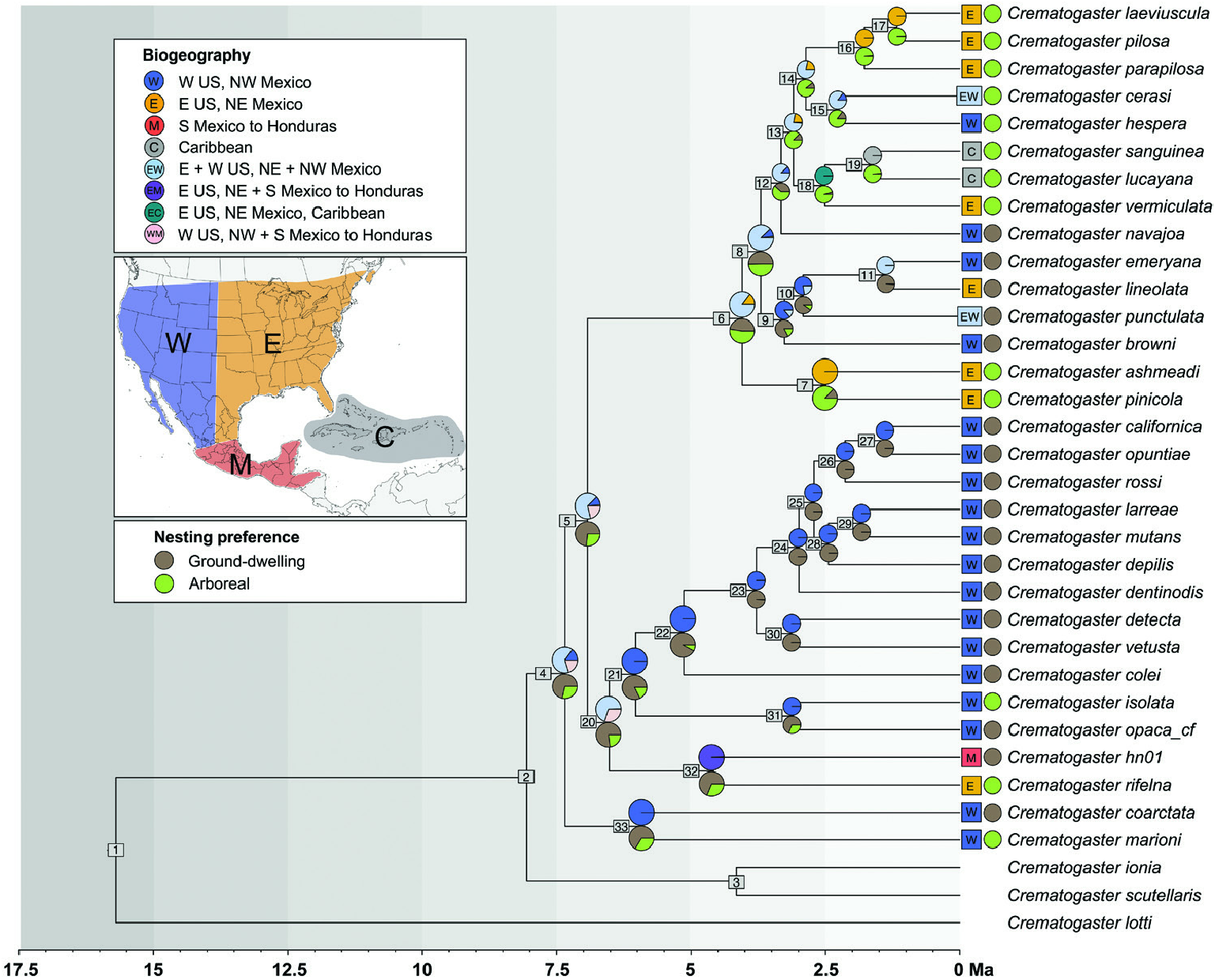
A single origin of the Nearctic diversity of the subgenus Crematogaster (Crematogaster) had already been inferred by Blaimer (2012a). This study had estimated a dispersal of the C. scutellaris View in CoL group to the Nearctic from the Palaearctic region sometime between 5.2–7.4 Mya. Our divergence analyses tighten this estimate somewhat, by inferring a slightly older median crown age of the Nearctic C. scutellaris View in CoL group with 7.35 Mya (node 4; Fig. 2 View Figure 2 and Table 1) and suggesting a dispersal to North America thus shortly after the divergence with the Palaearctic ancestor of the group around 8 Mya (node 2; Fig. 2 View Figure 2 and Table 1). Our results hence confirm the relatively recent, Late Miocene origin of the Nearctic C. scutellaris View in CoL group. However, the crown C. scutellaris View in CoL group node was calibrated with an age range taken from the earlier study ( Blaimer, 2012a), and we would, therefore, not expect large deviations from the previous estimates. The previous study ( Blaimer, 2012a) was not able to distinguish between a transatlantic dispersal or a terrestrial dispersal via the Bering Strait landbridge between the Palaearctic and Nearctic region. Both the previous and current age estimates are older than the earliest estimated opening of the Bering Strait around 5 Mya ( Gladenkov et al., 2002) and agree with a possible dispersal via the second Beringian landbridge (sensu Sanmartín et al., 2008). Our biogeographic reconstructions were equivocal with regard to the ancestral range of the most recent common ancestor of the C. scutellaris View in CoL group, and due to minimal sampling of Palaearctic species (as outgroups), we are also not able to pinpoint the exact source of the colonization event. Nonetheless, given that that the first split in the Nearctic C. scutellaris View in CoL group is between an exclusively western lineage, the C. coarctata View in CoL clade, and all other taxa, we suggest that a trans-Beringian dispersal from the eastern Palaearctic could be a likely scenario, as has been suggested for other ant taxa ( Jansen et al., 2010; Schär et al., 2018) and several groups of insects ( Sanmartín et al., 2008). More decisive evidence will have to await a larger biogeographic analysis including the Palaearctic members of the C. scutellaris View in CoL group and further outgroup sampling.
BIOGEOGRAPHY AND BIOLOGY OF NEARTIC CREMATOGASTER – FROM WESTERN GROUND- NESTERS TO EASTERN TREE- DWELLERS
Our phylogenomic analyses recovered well-resolved species-level phylogenies for the Nearctic C. scutellaris group. We found the group to be separated into three major clades: the small western USA and northwestern Mexico endemic C. coarctata clade, and the two species-rich C. opaca and C. lineolata clades, which have a predominantly western or eastern distribution, respectively. Interestingly, morphological characteristics to distinguish these three clades are lacking, at least in the workers, but perhaps this is not so surprising given their recent divergences (4–8 Mya; Fig. 2 View Figure 2 ; Table 1). The biogeography and diversification history of the Nearctic C. scutellaris group is thus characterized by a ‘western to eastern’ geographical pattern of range evolution, potentially dominated by the Rocky Mountain Range and the Great Plains as major dispersal barriers. Our range reconstructions are ambiguous about the early evolutionary history of the C. scutellaris group and show a broad eastern and western North American distribution (EW, nodes 4, 5 and 20; Fig. 2 View Figure 2 ). From these broad ancestral reconstructions, the subsequent limitation of Crematogaster lineages to a western or eastern distribution would have to be interpreted as range restrictions rather than dispersal events. However, since the C. coarctata subclade, which is sister to all other New World taxa, has a western North American distribution, it appears also probable that the MRCA of the entire Nearctic C. scutellaris group was distributed only in western North America during their early diversification.
Under the latter scenario, only one dispersal from western into eastern North America would have occurred within the C. opaca clade, when the ancestor of C. rifelna and an undescribed species migrated east sometime in the Late Miocene or Early Pliocene (branch leading to node 32; Fig. 2 View Figure 2 ), and then further south into Mexico and Mesoamerica. Considering the present distribution of these two sister-species in southeastern Texas and southern Mexico to Mesoamerica, this dispersal could have happened via migration through the low-elevation break between the Rocky Mountains/ Colorado Plateau and the Sierra Madre Occidental. Alternatively, a widespread MRCA for the C. opaca clade may have diverged from the western lineage as populations became isolated geographically and speciated. Judging by our phylogeny, it appears that most of the diversification of the C. opaca clade remained restricted to the western USA and northwest Mexico, west of the Great Plains. However, as we discussed above, several morphologically similar taxa occur south of the United States, in what we term the ‘ C. opaca complex’, which were not sampled here. Until the number of species and their phylogenetic affinities have been determined, the evolutionary history of the C. opaca clade cannot be fully resolved.
In contrast to the pattern seen in the C. opaca clade, there appears to have been a surprising amount of connectivity or exchange between biogeographic regions within the C. lineolata clade. If we accept a widespread Nearctic ancestor for this clade, as inferred in the present analysis and Fig. 2 View Figure 2 (node 6), we have to assume several subsequent range reductions – four to eastern North America (plus one dispersal to the Caribbean) and three to western North America. If we consider a possible western MRCA, five dispersals to the eastern realm and two range reductions from an EW to a W state would be necessary to explain current distribution patterns. We had recovered some phylogenetic incongruence between analyses in this clade, where some ML analyses placed C. laeviuscula , C. pilosa and C. parapilosa as sister to C. vermiculata , C. sanguinea and C. lucayana, instead of sister to C. hespera and C. cerasi , as in the main analysis. This is noteworthy as this topology would require fewer dispersal events or range reductions from western to eastern North America to explain the species distributions within this clade.
Regardless of this remaining uncertainty, we overall note a dominance of dispersals or range restrictions directed from the western Nearctic to the eastern Nearctic region, rather than vice versa. This is in line with previous findings. Sanmartín et al. (2008) analysed dispersal patterns across 57 animal clades in the Holarctic and also observed a statistically significant dominance of west to east dispersals. Several palaeoclimatic changes are discussed by these authors that could explain this trend ( Sanmartín et al., 2008), albeit not all apply to the time frame of our present study. Relevant for range evolution within the Nearctic C. scutellaris group may have been the increasing aridification of the central Great Plains in North America throughout the Pliocene ( Eronen et al., 2012). This process may have been an essential force in driving humid- and forest-adapted species eastwards, as mixed mesophytic forests shifted eastwards and were replaced by more xerophytic plant species ( Tiffney, 1985). Further climatic cooling and glaciation events during the Pleistocene probably reinforced this pattern, and the Great Plains are assumed to have continued to act as a dispersal barrier to the west for less aridadapted insects ( Howden, 1969). Among Nearctic members in the C. scutellaris group, only two species with a predominantly eastern distribution ( C. cerasi and C. punctulata ) traverse the Great Plains and marginally enter the west. It is also noteworthy that no western species penetrates the Great Plains to any significant degree.
The ancestor of North American C. scutellaris group is estimated to have a ground-dwelling habit, and this nesting preference was retained largely within the C. coarctata and C. opaca clades. Within members of the C. lineolata clade, the preference for grounddwelling is further retained largely in members with a western distribution. In fact, most western species are ground-dwellers, mainly nesting under stones, and only three species with a western distribution – one in each of the three main clades – are arboreal nesters: C. marioni , C. isolata and C. hespera . From the standpoint of biogeographic and climatic range evolution discussed above, the association of grounddwelling with the western range seems logical. The MRCA of the Nearctic C. scutellaris group must have been relatively cold-adapted to migrate across the trans-Beringian Bridge that was then covered by taiga ( Sanmartín et al., 2008). Climatic fluctuations during the Pliocene and Pleistocene would have then prevented, or at least disfavoured an invasion of the tree canopy as a nesting space by the ants. It is, therefore, plausible that the arboreal nesting habit was relatively recently acquired in the three western arboreal species. By contrast, all but one of the exclusively eastern or Caribbean species have an arboreal nesting preference. These arboreal inhabitants of mixed hardwood, subtropical forests and coastal scrublands may have survived the Pleistocene in glacial refugia, which occurred in eastern Texas/ western Louisiana and Florida ( Swenson & Howard, 2005). The exception, the ground-dwelling C. lineolata , is one of the most widely distributed species within the Nearctic C. scutellaris group, occuring from eastern Canada to the southern United States and west to the Great Plains. A generalist ground-nesting habit may have facilitated the success of this species, by possibly allowing a more widespread distribution even during glacial maxima or a more rapid post-glaciation range expansion.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Crematogaster undetermined
| Ward, Philip S. & Blaimer, Bonnie B. 2022 |
C. coarctata
| Mayr 1870 |