
Xeniamia atrithorax, Fraser, Thomas H. & Prokofiev, Artem M., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4144.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:5E49500D-AEEB-495A-9A2E-2BB0429CCB99 |
|
DOI |
https://doi.org/10.5281/zenodo.5680536 |
|
persistent identifier |
https://treatment.plazi.org/id/4235D83B-3CB9-40B8-A5F7-CDDCE9AA9457 |
|
taxon LSID |
lsid:zoobank.org:act:4235D83B-3CB9-40B8-A5F7-CDDCE9AA9457 |
|
treatment provided by |
Plazi |
|
scientific name |
Xeniamia atrithorax |
| status |
sp. nov. |
Xeniamia atrithorax new species
Figures 1–8 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 , Table 2 View TABLE 2
Diagnosis. A miniature species of Xeniamia ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ) attaining less than 30 mm SL with the following color pattern and morphologic characters: a large melanophore patch anterior to insertion of pelvic fin, extending forward on each side of isthmus, progressively thinning out ( Fig. 2 View FIGURE 2 ), black stomach, anterior intestine pale with black spots black intestine from second bend, interior of lower gill chamber blackish with concentrated melanophores extending forward to dorsal part of isthmus ( Fig. 3 View FIGURE 3 ), dorsal fins VIII-I,9 eighth dorsal spine visible, fourth dorsal spine longer than third, anal fin II,8, nine gill rakers.
Description. Holotype ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ): proportions as percentage of SL in Table 2 View TABLE 2 . All scales missing on head and body; dorsal fins VIII–I,9; anal fin II,8, damaged; pectoral fins 14–14; pelvic fins I,5; outer row of first gill arch with 2 rudiment, 1 raker on upper arch, and 2 rudiments and 8 rakers on lower arch, 6 pseudobranch filaments.
Villiform teeth in band on premaxilla & dentary; one row on vomer; one row (long) on palatine; none on basihyal or ectopterygoid.
Damaged preopercle lower edge with smooth, unossified flap, vertical bony edge smooth, bony ridge edge smooth.
Vertebrae 10+14, one ossified supraneural as /0/1/, two epurals, no paired uroneurals, no terminal bony sheath over third and fourth hypural, fused hypural 1+2, parhypural free, crest on preural centrum 2; principal caudal finray 17, upper and lower unbranched.
Preserved color pattern ( Figs 1 View FIGURE 1 & 2 View FIGURE 2 ): head with scattered melanophores dorsally from between eyes posteriorly to anterior edge of nape, a few melanophores in a patch near posttemporal; scattered melanophores at insertion of pelvic fins and on pale region posterior to a large melanophore patch extending forward on each side of urohyal, progressively thinning out; no melanophores on any fins or at fin-bases.
Holotype Paratypes Internal melanophore organization ( Figs. 3 View FIGURE 3 & 4 View FIGURE 4 ): Peritoneum silvery with speckled black spots becoming more dense in distribution ventrally, anterior wall black at connection with pharynx, black tissue extending forward along the floor of the oral cavity to below tongue; swim bladder whitish, no black spots; gonad pale with diffuse black spots; dorsal vein to sinus with black spots; intestine pale with black spots becoming more densely spotted past the first intestinal turn to completely black by the second intestinal turn thence black to anus along ventral portion of abdominal cavity; two large, pale pyloric caeca extending along the lower portion of the stomach; stomach black on upper half and with dense black spots on lower half; liver and heart pale.
Live color pattern unknown.
Gravid female, ova well-developed, extruding posteriorly.
Paratypes. Proportions in Table 2 View TABLE 2 . Pectoral-fin rays 14–15; lateral line scales 20–22 (8 specimens); circumpeduncular scales about 8; predorsal scales about 4; transverse row of scales above lateral line and first dorsal fin 3; transverse rows of scales below lateral line and anal fin 5 (transverse count based on one specimen); existing scales on body ctenoid except cycloid around paired-fin bases; and two cleared and stained specimens with one supraneural as /0/1/, fused hypurals 1+2, no terminal centrum sheath over the third fourth hypural, no evidence of paired uroneurals, 10+14 vertebrae; one cleared and stained specimen has a single epural ( Fig. 5 View FIGURE 5 ); radiographs of 36 specimens, one shown ( Fig. 6 View FIGURE 6 ). Many specimens have the first and/or eighth dorsal spines missing.
Cranial crests Y-shaped, well-developed, supraoccipital crest moderate, basisphenoid absent, otic bulla incompletely ossified.
Six infraorbitals ( Fig. 7 View FIGURE 7 ) with upper and lower edges smooth, lachrymal and second infraorbital expanded, third infraorbital without suborbital shelf; single large extrascapular; posttemporal edge smooth.
Maxilla with short expanded posterior plate; supramaxilla absent; junction between metapterygoid and hyomandibula interdigitated; seven branchiostegal rays (4 + 3); junction between ceratohyal and epihyal smooth; upper margin of ceratohyal deeply but smoothly concave without foramen; urohyal low and long, incised on posterior margin; opercular bones incompletely ossified; lowermost pectoral bony radial much larger than three upper ones, 2–1–1; pelvis with short and broad posterior processes, slightly divergent and broadly separated from each other medially; basiventral processes short (approximately equal in length to posterior processes), rod-like, strongly divergent; proximal-middle element of first anal-fin pterygiophore ending before first haemal spine, with 2/3 or 1/4 interdigitating pterygiophores; last pair of pleural ribs modified, expanded and shortened.
Many specimens with fin and body damage caused by trawls.
Otolith ( Fig. 8 View FIGURE 8 ). Oval (immature specimens) to sub-rhomboidal (adults), moderately deep otolith up to 2.5 mm length. Dorsal rim weakly convex, entire, with indistinct predorsal angle and strongly pronounced broad postdorsal angle. Ventral rim strongly convex, entire to indistinctly lobed; posterior rim strongly oblique, with more or less developed incision at junction with ventral rim; this incision is less developed in youngest specimens. Sulcus ostial, heterosulcoid; ostium and cauda of equal length; cauda narrow, straight bearing a distinct crest just above the crista inferior; colliculi heteromorphy; anterior colliculum oval, raised towards dorsal edge; posterior colliculum narrow, raised towards ventral edge; collum and pseudocolliculi absent; ostium possessing a conspicuous widening between anterior colliculum and crista superior; cristae superior and inferior moderately expressed. Rostrum large and massive, bluntly pointed, median; antirostrum moderately to well-developed, broad, rounded to bluntly pointed; excisura sharply angled. Dorsal depression large but shallow; ventral furrow distinct along whole length of ventral rim. Outer and inner surfaces weakly convex, slightly more on outer surface. Length:height ratio = 1.4–1.6; length:width ratio = 4.5–4.9.
Distribution. South China Sea off Khanh Hoa Province, Vietnam (Nha Trang and Van Phong Bays).
Etymology. Atrithorax from ater, atra, atrum Latin adjective meaning black and thorax Greek masculine noun meaning breastplate, the compound referring to the position of numerous melanophores as a dark spot in advanced of the pelvic-fin base, a noun in apposition.
Habitat. This species has been taken in shrimp trawls on muddy bottom. Most of specimens were collected from depths 70–119 m, but few samples were made at depths 40–60 m (currently in the unsorted part of Vietnamese collection in IOM and not listed here in the material examined) in the main waterway of the Bay of Nha Trang. The latter samples might have been pumped through canyons cutting out the outer shelf as proposed for other fish taxa collected from the unusually shallow depths in this area ( Nielsen & Prokofiev, 2010). There were no specimens or fragments of sea urchins commonly trawling together with Siphamia specimens. Year-to-year fluctuations in density of population were observed – this species was present at 18 of 22 stations trawling eastward and southward of Pyramid Island in 2005 and 2007 and two of 17 stations sampled in 2006 and 2009 –2012. All samples were made between April and July, and no specimens were collected during a winter monsoon season (October–January). This species always appeared in groups of about 10- 60 specimens per trawl, with many specimens having ripe ova or brooding eggs.
Reproduction. The smallest specimen, 14.5 mm SL was immature. Males with buccal enlargement suggestive of brooding eggs were 22.0–24.0 mm SL and showed loss of teeth on the vomer and palatines. A 23.0 mm SL male had approximately 280 orangish eggs, 0.5 mm in diameter. Females with developed ova were as small as 23.0 mm SL. The largest gravid female was 29.5 mm SL. No possible indications for commensal relationships of this species were present.
Remarks. The remarkable organization of melanophores raises questions about what these patterns might play in the life of this species: 1) suppressing the emission of light cause by luminous prey or 2) the use of chemicals partially from prey and reactions to create light in the pyloric caeca and intestine. Parsimony suggests that a single completely black alimentary canal or some variation of the length is all that is needed for the suppression of light. The complicate system of melanophore patterns and speckled anterior intestine coupled with well-developed pale caeca may allow for light to emanate from the anterior speckled isthmus region. It is not clear why the sinus vein or the artery below the swim bladder have melanophores. The dense melanophores forming a blackish spot in advance of the pelvic-fin base is unique among all known species of apogonids.
We use the long fourth dorsal spine as an indication of relationship with Jaydia rather than with the few species of Ostorhinchus that share this characteristic.
Gon (1997) revised species of Jaydia Smith (1961) as a subgenus of Apogon Lacepède (1801) with little osteological information, referring to Fraser (1972) who treated many species in the broad non-monophyletic subgenus Nectamia Jordan 1917 including five of those now in Jaydia . There are differing luminous systems described for Jaydia : 1) Weber (1909, 1913, plate 10 figure 7) and Weber & de Beaufort (1929) partially described for Jaydia argyrogaster ( Weber 1909) ; Gon & Allen (1998) a more completely described this unique system for Jaydia photogaster ( Gon & Allen 1998) . 2) Kato (1947), as Apogon marginatus (an unavailable name by Döderlein in Jordan & Snyder 1901, see Eschmeyer et al. (2016), now J. truncata ( Bleeker 1855) and Iwai & Asano (1958), as Apogon ellioti Day 1875 , now Jaydia smithi Kotthaus 1970 . A list of known and likely luminous apogonids is given in Table 1.
There are about 17–18 valid species of Jaydia (see Eschmeyer et al. 2016). Gon (1997) recognized ten species and nine synonyms. Nine more nominal species (three or four appear valid) were not treated in Gon’s revision. Three additional species have been described since Gon’s revision. We examined radiographs for about 11–12 valid species (see comparative material) of Jaydia yielded the following: all had three supraneurals as 0/0/0–1/, all had three epurals, all had five free hypurals. An eighth dorsal spine was reported for Jaydia erythrophthalma by Gon et al. (2015) but provided limited information about other osteological characters. Radiographs confirm the 3 supraneurals described by Gon et al. and we observed three epurals, five free hypurals. All other Jaydia have seven visible first dorsal-fin spines.
The otoliths of Apogonidae are not sufficiently known, though preliminary data based on study of the various Vietnamese taxa indicate morphologic diversity in the sagittal structures. Xeniamia shares a marked dorsal widening of ostium and a narrow cauda bearing a collicular crest, both of which are characteristic for the apogonid sagittae, but the poorly developed predorsal angle and a conspicuous excision at posterior ventral margin appear to be peculiar for the new taxon. All small specimens less than 23 mm SL examined have otoliths strongly corroded (by formalin fixation) and partially disintegrated. However, it is clear, that immature otoliths are more oval in shape, with feebly developed ventroposterior incision. Variations in shape of otolith in adult specimens close to maximal known size (SL 27.0– 28.5 mm) are considerable, particularly in shape of dorsal and ventral rim, rostrum and antirostrum and in expression of ventroposterior excision. Three otoliths shown in Figure 7 View FIGURE 7 look like growth changes rather than individual variations; however, they were taken from fishes of similar size, all of which are females with ripe eggs close to spawning. Together with poor ossification of some skeletal elements this may suggests a neotenic nature of Xeniamia .
Material examined. Holotype USNM 436743, 25.1 mm SL, female, Van Phong Bay ,12°40.944'– 12°36.924'N 109°29.752' – 109°30.002'E, Vietnam, 30 May–1 Jun 2007, 97– 119 m, 6 m shrimp trawl, 2247–0039 hrs, A.M. Prokofiev, x-ray.
Paratypes: CAS 23808 (2, 21.5–24.3), ROM 98985 (2, 25.1–27.2), UF 237134 (3, 20.4–24.1) & USNM 436744 About USNM (2, 20.5–25.5), all same data as the holotype, 5 specimens x-ray . ZMMU 23156 View Materials (20, 22.0–29.5) all same data as the holotype, x-ray, (2, 24.5–25.0) cleared & stained . ZMMU 23157 View Materials (19, 14.5–29.0), Van Phong Bay , 12°35.044'– 12°29.927'N 109°30.004' – 109°30.018'E, Vietnam, 30 May–1 Jun 2007, 95– 120 m, 6 m shrimp trawl, 0144–0402 hrs, x-ray, A.M. Prokofiev . ZMMU 23158 View Materials (14, 16.0–29.0), Nha Trang Bay , off Hon Dun (Pyramid Island), 12°13.200'– 12°11.329'N 109°24.161' – 109°23.944'E, Vietnam, 21 May 2005; 70-80 m, 6 m shrimp trawl , 2045–2145 hrs, x-ray, D.A. Astakhov & A.M. Prokofiev.
Comparative material. All in Jaydia with holotypes and syntypes listed by original genus, alphabetical by species: Amia albomarginata Holotype, USNM 68402, 83.6 mm SL, Cavite, Luzon I., Philippines, x-ray. Apogon argyrogaster Syntypes , ZMA 101.075, 2 , 34.9–47.5 mm SL, near western coast of New Guinea , Siboga Expedition. Jaydia carinata USNM 175735, 1 , 97.0 mm SL, Marinduque I., Philippines, Albatross 3009, 2 Mar 1909, x-ray . USNM 71527, 5 , 81.8–94.4 mm SL, Shimizu Suruga , Japan, Albatross , 1906, x-ray. Apogon ellioti Lectotype, ZSI F1904, 77.5, Madras , India, x-ray. AMS Paralectotype, B8226, 77.3, Madras , India, x-ray. Jaydia erythrophthalma Paratypes CAS 236504, 2 , 50.3–57.0 between Luzon & Mindoro, Philippines, 1 June 2011, 115 – 144 m, x-ray. Apogon fuscomaculatus Holotype, NTM S. 13284–014, 48.2 mm SL, NE of Charles Point , Beagle Gulf, Northern Territory, Australia, 2 Sep 1992, 18 – 24 m, trawl, photograph, x-ray. Mionorus heraldi Holotype, SU 38263, 104.5 mm SL, Ragay Gulf, Luzon , Philippines, x-ray. Jaydia hungi SAIAB 3116, 2 , 87.6–96.5 Comoro Ids., Jun 1959, x-ray . SAIAB 3117, 3 , 88.2–100, Pemba I., Tanzania, x-ray. Jaydia lineata USNM 71108, 4 , 64.3– 709, Nanao, Honshu I., Japan, x-ray. Jaydia novaeguinae USNM 177675, 2 , 53.8–55.9 Peitaiho, China, x-ray . USNM 51965 About USNM , Southern Negros, Philippines, online x-ray . USNM 163229 About USNM , Cavite, Luzon, online x-ray Apogon photogaster Holotype WAM P.31213–057, 49.0 mm SL, Padoz Reef , Madang Lagoon, Papua New Guinea 21 Oct 1996, 18 – 23 m. Paratype USNM 348213, 43.6 mm SL, same data as holotype, photograph, x-ray . Paratype BPBM 38301 About BPBM , 42.0 mm SL same data as holotype, x-ray . Paratype WAM P. 30358–001, 29.8 mm SL, Madang Lagoon, Padoz Reef , 28 Oct 1991, 26– 30 m, x-ray . Paratype SAIAB ( RUSI) 57597, 47.4 mm SL, Madang Lagoon, Padoz Reef , 28 Oct 1991, 26– 30 m, cleared & stained, data via Ofer Gon . BPBM 32628, 1 , 49.0 mm SL, Tripod Reef, lagoon off of Nagada Harbor, Papua New Guinea, 18 Nov 1987, 30 m, photograph, x-ray. Apogon quartus Holotype , USNM 307688, 49.8 mm SL, Saya de Malha Bank, 11°05'00"S 62°02'00"E, x-ray. Apogon queketti Syntype, SAM 11657 1, 75.1, Natal , South Africa, 40 fms . Syntypes, SAM 11658 5, 44.1–77.4, Natal , South Africa, 40 fms . SAIAB ( RUSI) 3177, 2, 64.4–68.5, Durban, Natal, South Africa. Jaydia smithi Paratypes , ZMH 5034 View Materials (ex IOES 120 B,E, F), 5, 12.7–46.3, Gulf of Aden, 11°17.9'N 49° 01.3E, Meteor Sta 97, 42 m, 18 Dec 1964, xray GoogleMaps . USNM 149365, 4 , Manila Bay, Philippines, online x-ray. Amia striata Holotype , USNM 68403, 69.8 mm SL, Manila Bay/ Lingayen Gulf, Luzon, Philippines, D 5442, 82 m, x-ray. Paratypes , USNM 93410 About USNM 11, 31.5–66.6, same data as holotype, x-ray. Apogon tchefouensis Paratype , MNHN 41–148 About MNHN , 46.0, China, x-ray . Paratype, MNHN 41– 149 About MNHN 36.2 mm SL, China, damaged, x-ray. Jaydia truncata USNM 71275, 5 , 80.4–86.3, Kagoshima, Satsuma , Kyushu I., Japan, x-ray . USNM 59635, 2 , 50.9–52.2, Matsushima Bay , Japan, x-ray . USNM 59636 About USNM , Yamagawa, Japan, online x-ray.



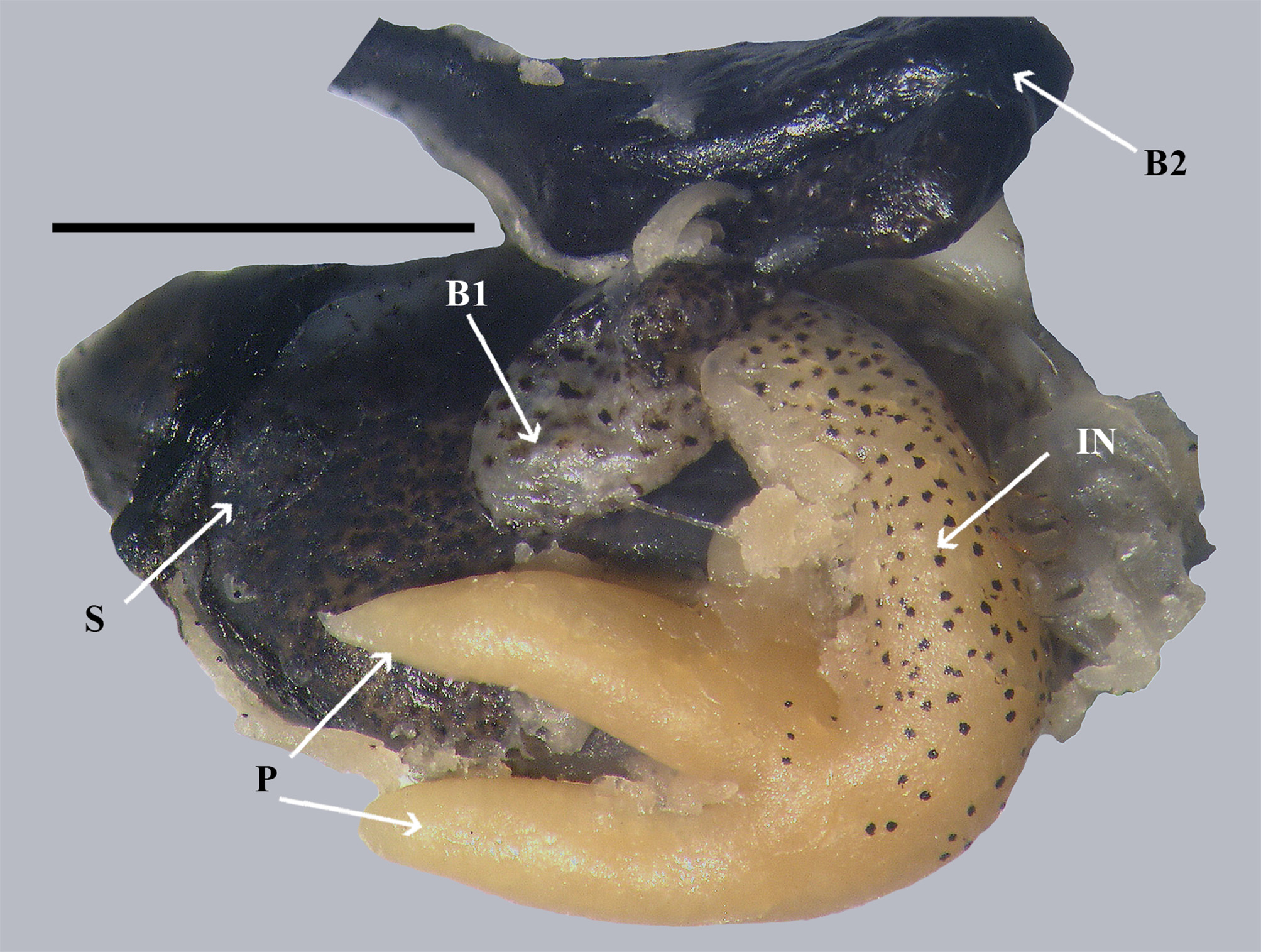

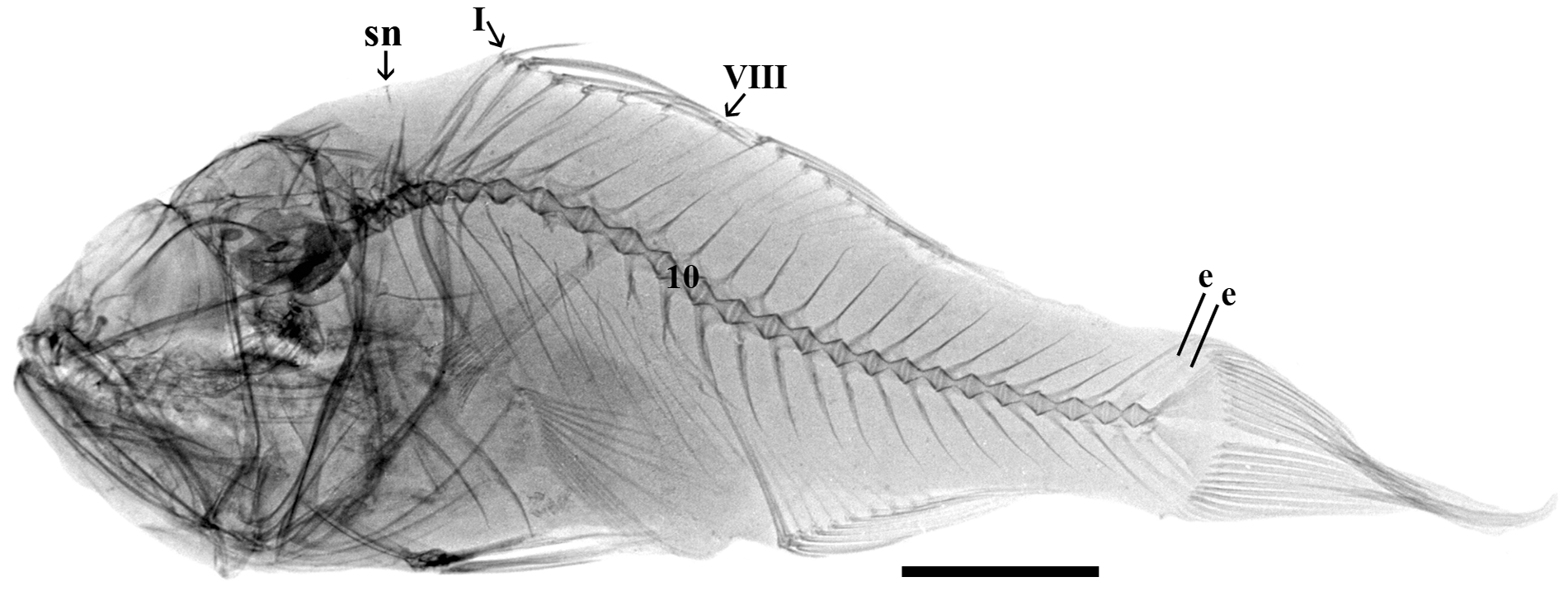
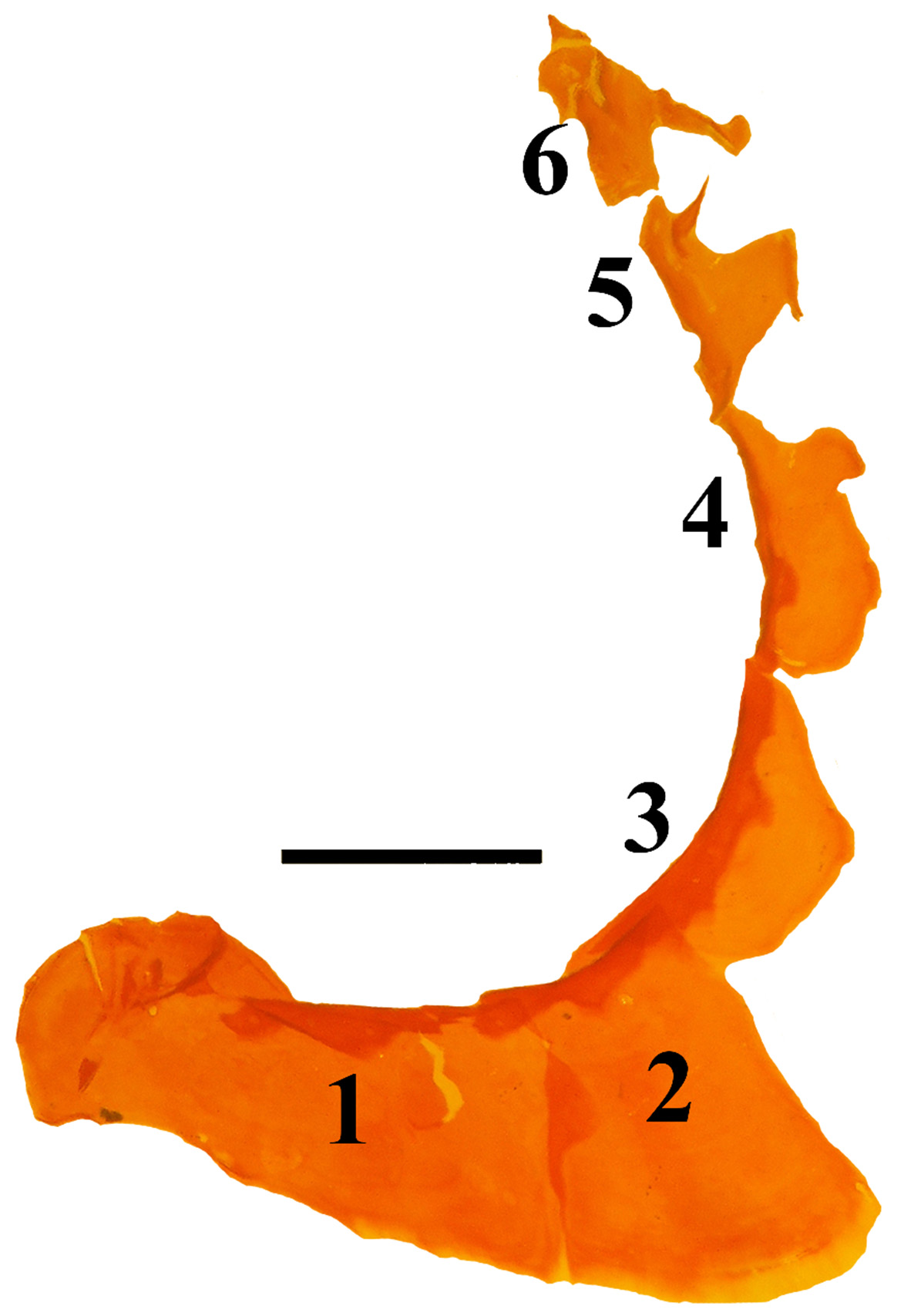

TABLE 2. Proportions as a percentage of standard length for the holotype of Xeniamia atrithorax with proportion ranges for two subsamples of all of the paratypes.
| USNM 436743 | ZMMU 23156 | ZMMU 23157 |
|---|---|---|
| 25.1 mm SL | Nha Trang Bay N=8 22.5–29.5 mm SL | Van Phong Bay N=10 22.0–29.0 mm SL |
| Percentages | ||
| Body depth 38.7 Head length 37 Eye diameter 8.76 | 38.0–40.4 38.9–42.0 10.0–10.2 | 35.7–43.2 37.9–44.4 9.26–11.4 |
| Snout length 5.98 Interorbit width 9.16 Upper jaw 22.3 | 7.20–9.62 8.93–10.0 20.0–23.0 | 7.72–11.4 9.26–11.6 21.1–25.0 |
| First dorsal spine 1.2 Second dorsal spine 6.77 Third dorsal spine 15.9 | 1.48–2.69* 4.80–10.4 12.5–14.2 | 1.11–3.46* 7.56–13.5* 12.8–18.2* |
| Fourth dorsal spine 16.7 Second dorsal fin spine. First anal spine 3.57 | 14.0–16.4 17.3–26.0* 3.05–4.80 | 14.8–18.2* 20.4–25.5* 2.76–4.56 |
| Second anal spine 9.16 Pectoral fin length. Pelvic fin length. | 8.47–10.4 25.0–30.0* 20.0–23.0 | 9.33–11.4 24.6–28.9* 20.7–24.1* |
| Caudal peduncle length 21.1 Caudal peduncle depth 13.2 | 20.0–23.1 12.0–14.3 | 19.0–25.0 13.1–14.9 |
| * = one or more element broken. = missing measurement because element broken |
| IOM |
Institute of Oceanology, Academy of Sciences |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
| CAS |
California Academy of Sciences |
| NTM |
Northern Territory Museum of Arts and Sciences |
| SAIAB |
South African Institute for Aquatic Biodiversity |
| WAM |
Western Australian Museum |
| BPBM |
Bishop Museum |
| RUSI |
J.L.B. Smith Institute of Ichthyology (formerly of Rhodes University) |
| SAM |
South African Museum |
| ZMH |
Zoologisches Museum Hamburg |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |