
Xanthorhoini
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3721.6.3 |
|
publication LSID |
lsid:zoobank.org:pub:3965D795-99D0-483A-BD05-2360913D0668 |
|
DOI |
https://doi.org/10.5281/zenodo.6163799 |
|
persistent identifier |
https://treatment.plazi.org/id/039287BC-FFB2-6048-1BC4-7CC44FEBFBA5 |
|
treatment provided by |
Plazi |
|
scientific name |
Xanthorhoini |
| status |
|
Tribe Xanthorhoini
Palaearctic species
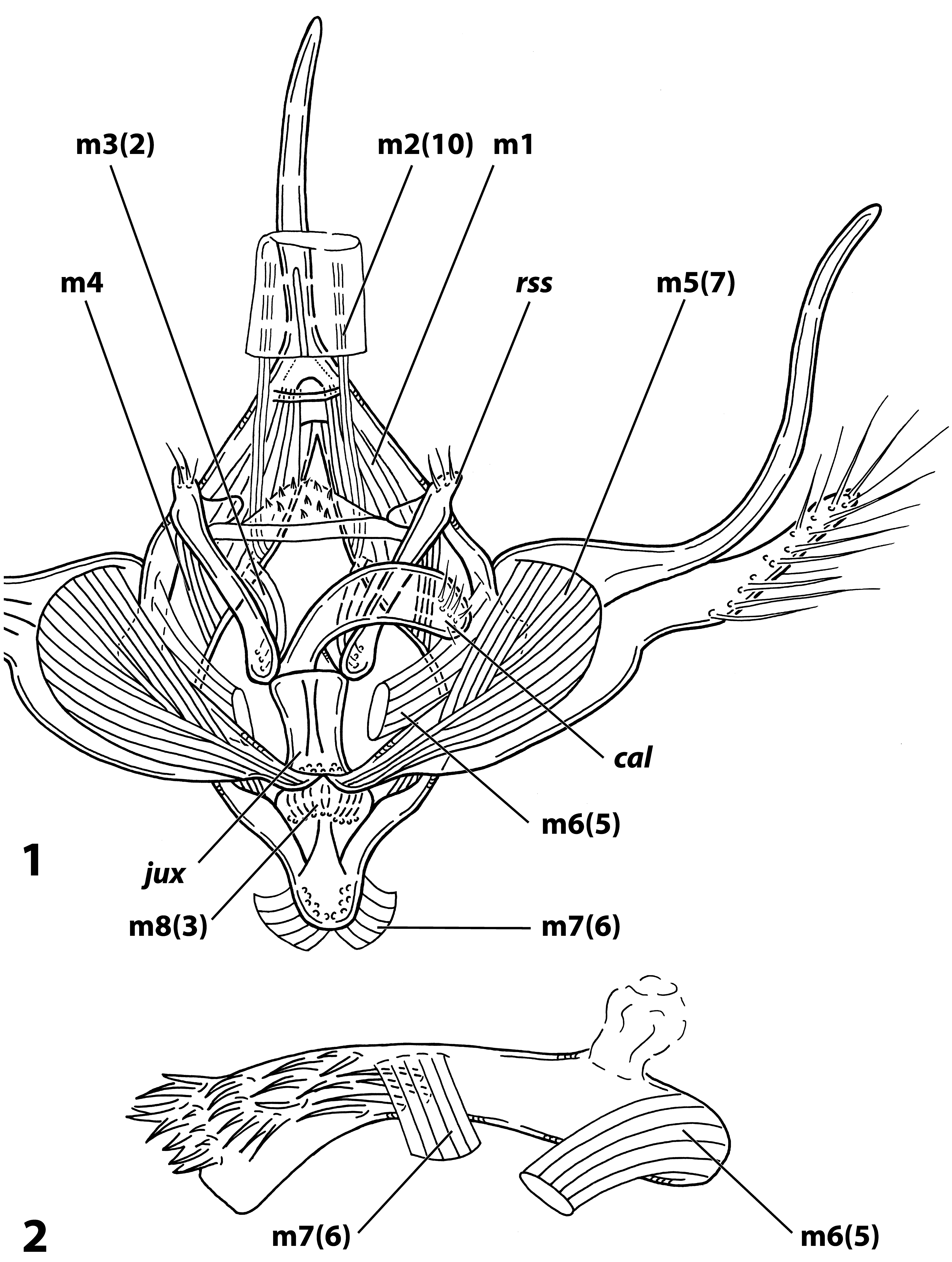
Xanthorhoe montanata (Denis et Schiffermüller) ( Figs 1–2 View FIGURES 1 – 2 )
The musculature of the male genitalia has been described by Razowskj and Wojtusiak (1981) and Valersky (2011). An important character of X. montanata and generally of the tribe Xanthorhoini s. str. appears to be the absence of outgrowth of the costal margin of valvae and accordingly absence of the muscles attached to this region. The tergal extensors of valvae, muscles m3(2) arise from the anterior margin of the tegumen and extend to the base of the rodshaped sclerites fused with the juxta laterally, calcar (=medial projection) is present. The tergal flexors of valvae (m4) connect the ventral parts of the anterior margin of the tegumen with the distal end of the rod-shaped sclerites.
Xanthorhoe designata rectantemediana (Wehrli) ( Figs 3–4 View FIGURES 3 – 4 )
The genital musculature of X. designata is very similar to X. montanata . The depressors of the uncus (muscles m1), the retractors of the anal tube (m2(10)) and the extensors and flexors of the valvae (muscles m3(2), m4, and m8(3)) have the same location as in X. montanata . The distinct suture between the tegumen and the vinculum is missing. The valvae are broader, costa is not produced into a long spine distally but the position of the intervalval muscles m(5)7 is the same. These extend from the central part of the valva to the sacculus, with a small bundle attached to the vinculum. The juxtal plate is elongated dorso-ventrally, and the calcar and rod-shaped sclerites are present. The aedeagus is very large, with broad protractors m6(5) attached to its basal process and extending to the medial part of the vinculum. A thin medial process of the elongated saccus is present, the muscles m7(6) run from the thin medial process to the medial portion of the aedeagus.
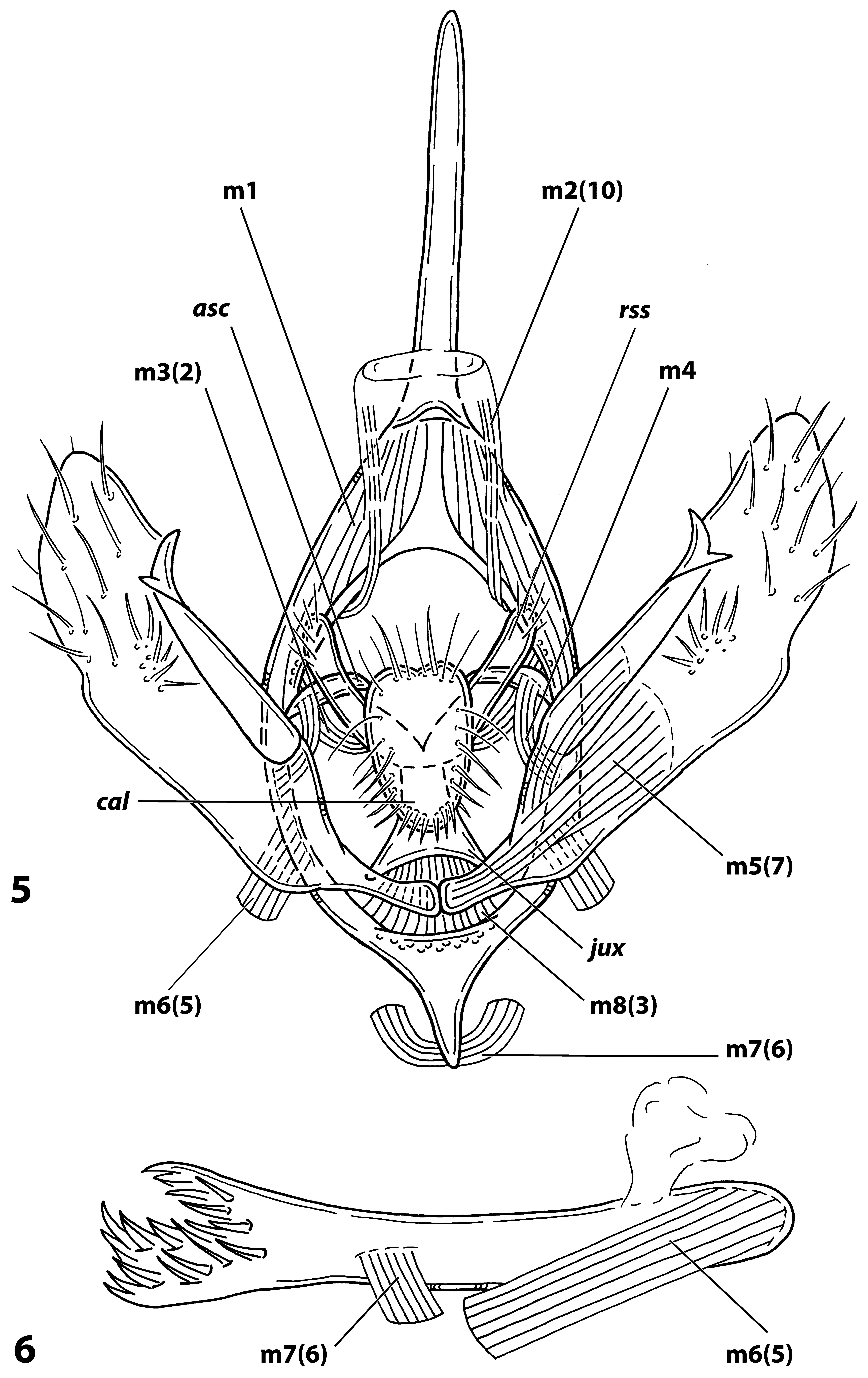
Xanthorhoe biriviata angularia (Leech) ( Figs 5–6 View FIGURES 5 – 6 )
The location of the muscles m1, m2(10), m3(2), m(5)7 and m8(3), as well as of the aedeagal musculature (m6(5) and m7(6)) is similar to that of the type species of the genus. A characteristic feature is the attachment of tergal flexors of the valvae (muscles m4). Although the juxtal rod-shape sclerites are present, like in X. montanata , the muscles in their distal part are missing. The muscles m4 run from the anterior margin of the tegumen to an arched sclerite between the tegumen and vinculum. The arched sclerite is not connected to the basal part of the costa valvae.
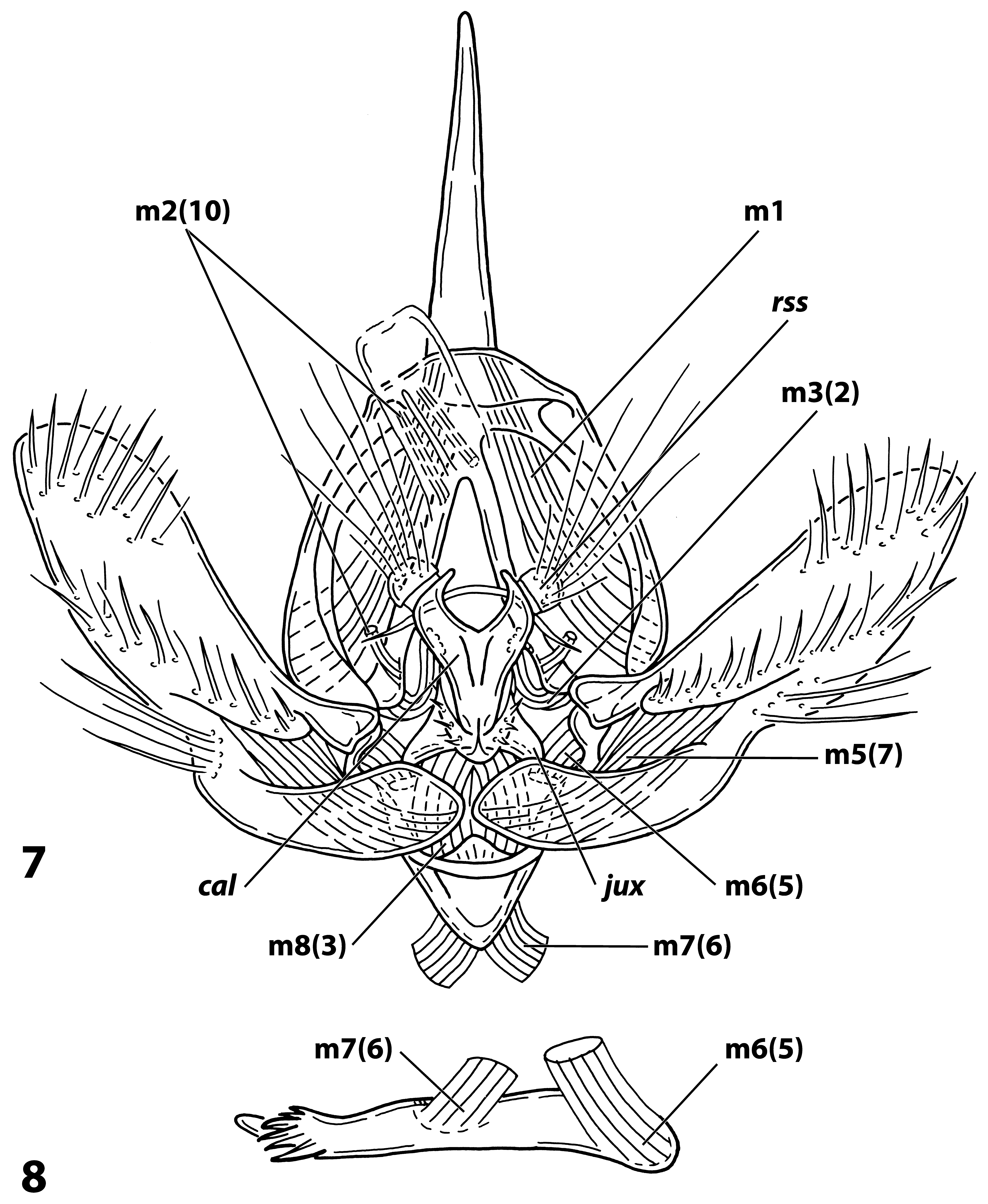
Camptogramma bilineata (Linnaeus) ( Figs 7–8 View FIGURES 7 – 8 )
Well developed muscles m1 connect the anterior margin of the strongly divided cupola-shaped tegumen with the base of the stout, medium-sized uncus. Two pairs of muscles are attached to the processes of the anterior margin of tegumen: the retractors of the anal tube m2(10) which run to the inner surface of the anal tube distally, and the muscles m3(2) which run to the membrane dorsad of the posterior part of compound juxta. The juxta is a fusion of the juxtal plate, the medial calcar and the lateral modified rod-shaped sclerites that are much reduced and membranously connected with the diaphragm. The muscles m8(3) extend from the anterior margin of the juxta to the posterior margin of the saccus. The muscles m4 are reduced due to modifications and reductions in the juxtal area. A fairly clear suture in the zone of articulation of the tegumen with the narrowed vinculum is present. The costa of a rather simple-shaped valva is weakly sclerotized. The muscles m5(7) consist of two bundles, one of them runs to the sacculus, whereas the second one runs to the middle part of the vinculum. The muscles m6(5) connect the keel of the basal process of the small aedeagus and the dorsal part of the vinculum close to the zone of its articulation with the tegumen. The muscles m7(6) extend from the medial portion of the aedeagus and run to the anterior margin of the saccus.
Epirrhoe alternata (Müller) ( Figs 9–10 View FIGURES 9 – 10 )
The uncus is medium sized and separated from the tegumen. The muscles m1 extend from the anterior margin of the tegumen to the base of uncus. The retractors of the anal tube m2(10) connect the tegumen with the inner wall of the anal tube distally. Broad muscles m5(7) are attached to the middle portion of the valva, ventrad of the flapshaped distal rounded enlargement and run to the sacculus, with a small bundle shifted to the vinculum. The calcar is present, flat, elongate heart-shaped, with hook-shaped medial projection proximally. The muscles m8(3) are unpaired, connecting the inner wall of the juxta medially to the posterior margin of saccus. The position of aedeagal muscles m6(5) and m7(6) corresponds to that in Xanthorhoe . The tergal flexors of valvae m4 were not found. The position of the muscles m3(2) is somewhat unusual for the tribe. A short horn-shaped sclerite is between the basal part of costa valva and the juxta. This sclerite seems to be homologous to a rod-shaped sclerite in other xanthorhoines although not completely shifted to the juxtal region. The connection of this sclerite to the basal part of costa valva is maintained through a minute sclerotized suture. Faint connection to the juxta is also present. The tergal extensors of valvae m3(2) are attached to a small protuberance on the tegumen under m2(10) and extend to the ventral part of the short horn-shaped sclerite.
Costaconvexa polygrammata (Borkhausen) ( Figs 11–12 View FIGURES 11 – 12 )
The male genitalia are heavily sclerotized. The tegumen is almost completely divided medially. The uncus and its depressors muscles m1 are enlarged. The position of m1 and m2(10) corresponds to that in X. montanata . The valval costa is sclerotized. Long muscles m5(7) extend from a broad area ventrad of the costa and run into the big sack-shaped sacculi. The muscles m8(3) connect the anterior margin of the juxta to the inner wall of the prominence of the posterior margin of a weakly pronounced saccus. A large complex of characters in the male genitalia of C. polygrammata appears to be autapomorphic. A conspicuous suture between the tegumen and the vinculum is present. A small divergent juxta bears a somewhat reduced, modified calcar, which is connected by membrane to the diaphragm and is seemingly homologous to the calcar of X. montanata . The rod-shaped sclerites are not developed. Rather large muscles m3(2) connect the anterior margin of the tegumen with the juxta, which is characteristic for the Xanthorhoini . The muscles m4 are reduced due to modifications and reductions in the juxtal area. The retractors of the aedeagus are massive and consist of two bundles. The larger one (m7a(6a)) extends broadly from the middle portion of the aedeagus, runs to the inner wall of the posterior margin of a weakly pronounced saccus into which it is inserted above a thin sclerotized suture. The smaller bundle (m7b(6b)) is attached to the basal process of the aedeagus and runs to the anterior margin of the saccus. The muscles m6(5) are inserted on the vinculum directly under the suture between the vinculum and the tegumen, the second place of attachment is the basal process of the aedeagus.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Larentiinae |
|
Genus |