
Typton anaramosae, Neves, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4768.2.7 |
|
publication LSID |
urn:lsid:zoobank.org:pub:0F2EE17B-8206-4070-BFF5-DB13FA147B58 |
|
DOI |
https://doi.org/10.5281/zenodo.3795093 |
|
persistent identifier |
https://treatment.plazi.org/id/193CE50B-2599-44D3-AD7E-9EB9BB046E42 |
|
taxon LSID |
lsid:zoobank.org:act:193CE50B-2599-44D3-AD7E-9EB9BB046E42 |
|
treatment provided by |
Plazi |
|
scientific name |
Typton anaramosae |
| status |
sp. nov. |
Typton anaramosae View in CoL sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
urn:lsid:zoobank.org:act:193CE50B-2599-44D3-AD7E-9EB9BB046E42
Type material. Cabo Verde Archipelago. Holotype: Ovigerous female (pocl 4.2 mm) [ MNCN 20.04/12401], São Vicente Island, Porto Grande Bay, Enseada de Corais da Matiota, 16°53’47.4”N 24°59’32.9”W, snorkeling, depth: 2–3 m, in sponge growing under large rock, leg. K. Neves, September 2019. Paratypes: male (pocl 3.4 mm) [ MNCN 20.04/12402] and ovigerous female (damaged and lacking second major cheliped, pocl not measured) [ UCV 2020/00016], same collection data as for holotype. Two ovigerous females (one: pocl 3.8 mm, other: damaged, pocl not measured), São Vicente Island, Porto Grande Bay, Enseada de Corais da Matiota, 16°53’47.4”N 24°59’32.9”W, snorkeling, depth: 2–3 m, in sponge growing under large rock, leg. K. Neves, 29 December 2019 [ UCV 2020/00017].
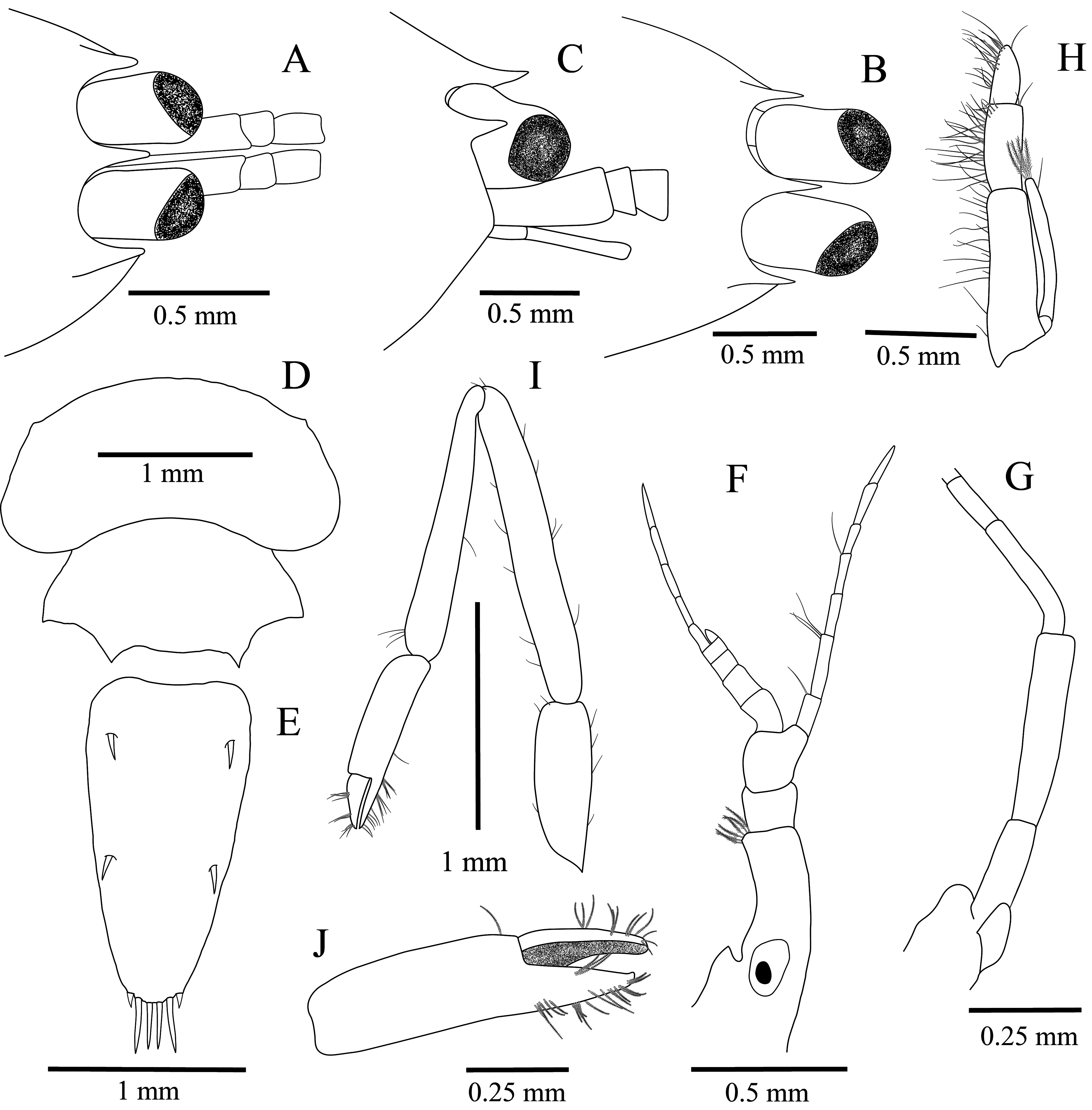
Description. Carapace ( Fig. 1 View FIGURE 1 A–C) subglobular, smooth, glabrous, slightly depressed, longer than deep. Rostrum ( Fig. 1 View FIGURE 1 A–C) short, triangular in dorsal view, unarmed, 0.67 length of ocular peduncle, tapering distally, acute, tip curved upwards in lateral view, ventral border convex throughout. Paraorbital processes ( Fig. 1 View FIGURE 1 A–C) well developed, 0.85 length of rostrum, slightly divergent in dorsal view, almost reaching base of cornea and pointing slightly upward in lateral view, reaching level of most-anterior pterygostomial margin. Orbit ( Fig. 1B View FIGURE 1 ) feebly demarcated, without distinct inferior orbital angle or antennal spine; pterygostomial margin slightly produced anteriorly, somewhat angular.
Eyes ( Fig. 1 View FIGURE 1 A–C) well developed, cornea rounded, set obliquely on stalk; stalk stout, subcylindrical, inner margin straight, longer than wide and subequal to diameter of cornea; anterior margin of cornea reaching distal third of first article of antennular peduncle.
Pleon elongate, smooth, glabrous, moderately expanded in ovigerous females, enclosing large number of small eggs; first 4 pleonites with pleura broadly rounded, 5th pleuron ( Fig. 1D View FIGURE 1 ) slightly produced, posterior margin of 6th pleonite without dorsal medial tooth, posterolateral and posteroventral angles acutely produced ( Fig. 1D View FIGURE 1 ).
Telson ( Fig. 1E View FIGURE 1 ) moderately slender, tapering, 1.9 times of 6th pleonite length, about twice as long as maximal width; lateral margins convex, posteriorly convergent; posterior margin feebly convex, without median process; dorsal surface with 2 pairs of spiniform setae, anterior pair 0.08 times telson length, situated at 0.2 of telson length, posterior pair similar in size, situated at about 0.6 of telson length; posterior margin with 3 pairs of non-setulose setae, lateral pair minute, 0.6 times as long as dorsal spines, intermediate pair long, slender, 0.17 telson length, median pair slightly shorter than intermediate pair.
Antennular peduncle ( Fig. 1F View FIGURE 1 ) with proximal article subcylindrical, 3.9 times longer than distal width, slightly broadened proximally; statocyst well developed, with statolith; stylocerite very short, acute, broad; intermediate article shorter than distal article, their combined length equal to about 0.4 length of proximal article; upper flagellum biramous, short, stout, four proximal articles of rami fused, short ramus with single short rounded subdivision, longer ramus short, slender, with 5–6 subdivisions; about 6–8 groups of aesthetascs; lower flagellum slender, short, with 7 subdivisions.
Antenna ( Fig. 1G View FIGURE 1 ) with stout unarmed basicerite bearing conspicuous antennal gland tubercle mesially; carpocerite slender, subcylindrical, about 5 times longer than distal width, 1.6 merocerite length; flagellum slender, short, 3.9 times as long as carpocerite; scaphocerite strongly reduced, 0.4 carpocerite length, 2.5 times as long as proximal width, tapering distally, terminally rounded, without distolateral spine or setae.
Mouthparts typical for genus, without species-specific features. Third maxilliped ( Fig. 1H View FIGURE 1 ) with antepenultimate article broad; penultimate article about half as long as antepenultimate article, 1.4 length of ultimate article; ultimate article slender, tapering towards apex; exopod reaching slightly beyond distal margin of antepenultimate article, with 1 short, simple, subdistal seta, in addition to 4 slender, plumose, distal setae.
First pereiopods ( Fig. 1 View FIGURE 1 I–J) exceeding antennular peduncle by distal third of merus. Palm of chela slender, subcylindrical, slightly compressed, about 3 times as long as wide. Fingers robust, short, stout, 0.45 of palm length, strongly spatulate; finger cutting edges entire, blade-like, unarmed; dactylus distally rounded in dorsal view, fixed finger similar to dactylus, tip feebly bidentate; outer and mesial surfaces of fingers bearing some tufts of serrulate setae. Carpus about 6.6 times longer than distal width, 2.2 times as long as palm, tapering proximally. Merus slen- der, about 7.4 times longer than broad, 1.1 times as long as carpus. Ischium short, stout, broader than merus, about 2.7 times as long as broad, 0.6 times merus length. Basis shorter and slenderer than ischium.
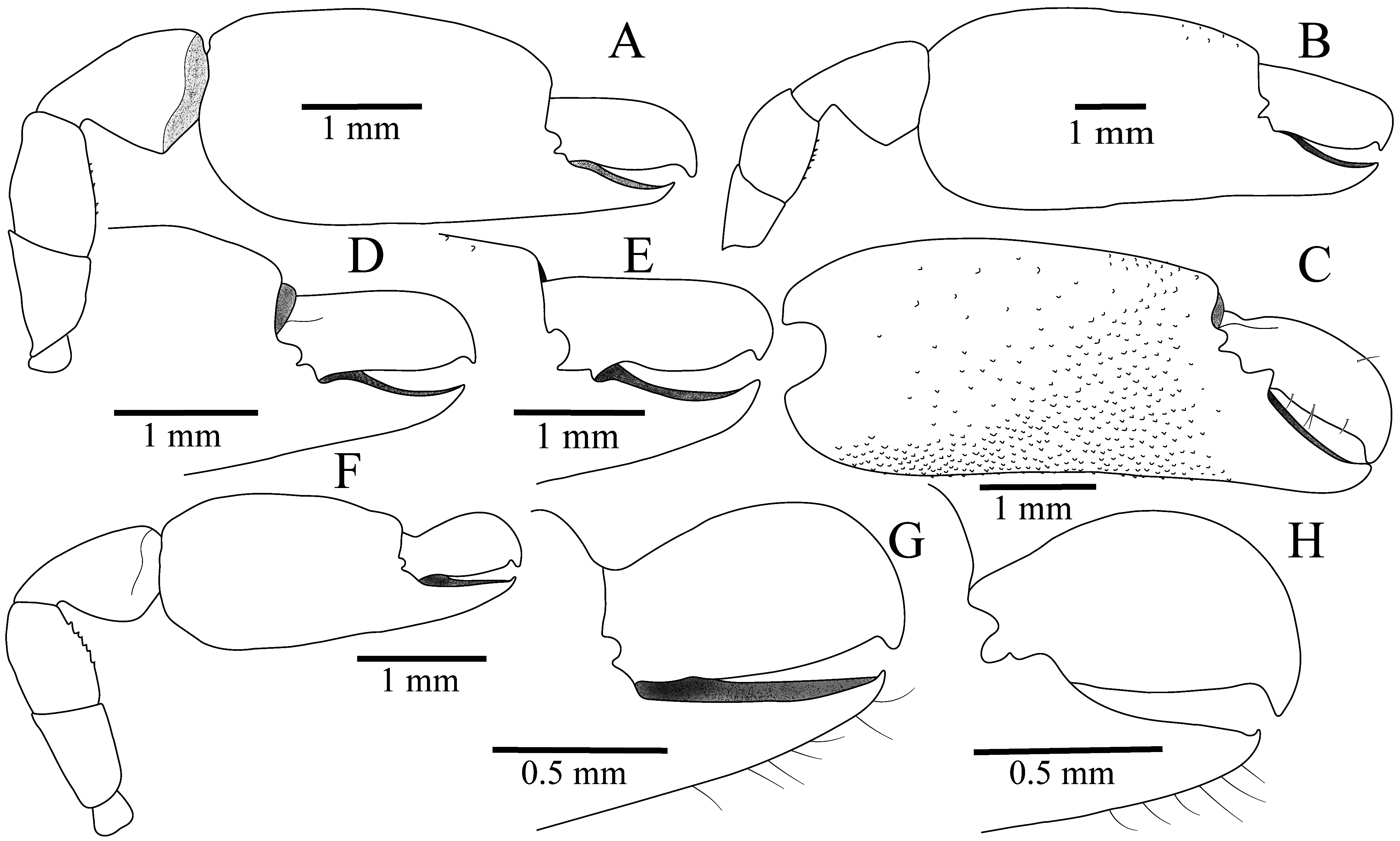
Second pereiopods well developed, unequal in size, dissimilar in shape, reaching beyond the antennal peduncle by entire carpal length. Major second pereiopod ( Fig. 2 View FIGURE 2 A–E) greatly enlarged, longer and more robust than minor second pereiopod; chela greatly enlarged, moderately swollen; palm bearing several small granules on mesial surface, extending to ventral and dorsal margins, slightly compressed laterally, sub-oval in cross-section, flattened medially, slightly tapering distally, length about 1.7 maximal height; dactylus laterally compressed, 0.34–0.37 times length of palm, with stout, corneous, bluntly hooked tip; dactylar cutting edge thickened, entire, unarmed; pollex about as long as dactylus, with curved, distally sub-acute, corneous tip; cutting surface opposed to dactylus with deep groove accommodating dactylar cutting edge; mesial edge of pollex with single, strong, blunt tooth proximally, lateral margin unarmed; both fingers curved mesially (inwards) at their tips; carpus 0.4–0.5 length of palm, ventral surface irregular, without distinct teeth, about 1.4 times as long as maximal width, narrow proximally, distally widening; merus 1.4 times as long as broad, 0.77 length of carpus and 0.37 length of palm, ventral surface with small, conical, sharp teeth; ischium 0.7 length of merus, unarmed.
Minor second pereiopod ( Fig. 2 View FIGURE 2 F–H) smaller than major second pereiopod; chela 0.68 of major chela length; palm suboval in cross-section, 1.7 times as long as maximal height, bearing sparse granules on mesial surface; dactylus 0.4 palm length, compressed, with curved tip; cutting edge sharp, entire; pollex excavated on surface opposed to dactylus; lateral edge sharp, entire; mesial edge projecting proximally as blunt triangular tooth, fingertips sharp, curved; carpus about half-length of palm, with minute conical teeth on ventral surface, 1.1 times maximal height, much wider distally; merus 1.4 times as long as broad, about 0.8 of carpal length and 0.41 of palm length, with minute conical teeth on ventral surface; ischium about as long as merus, unarmed.
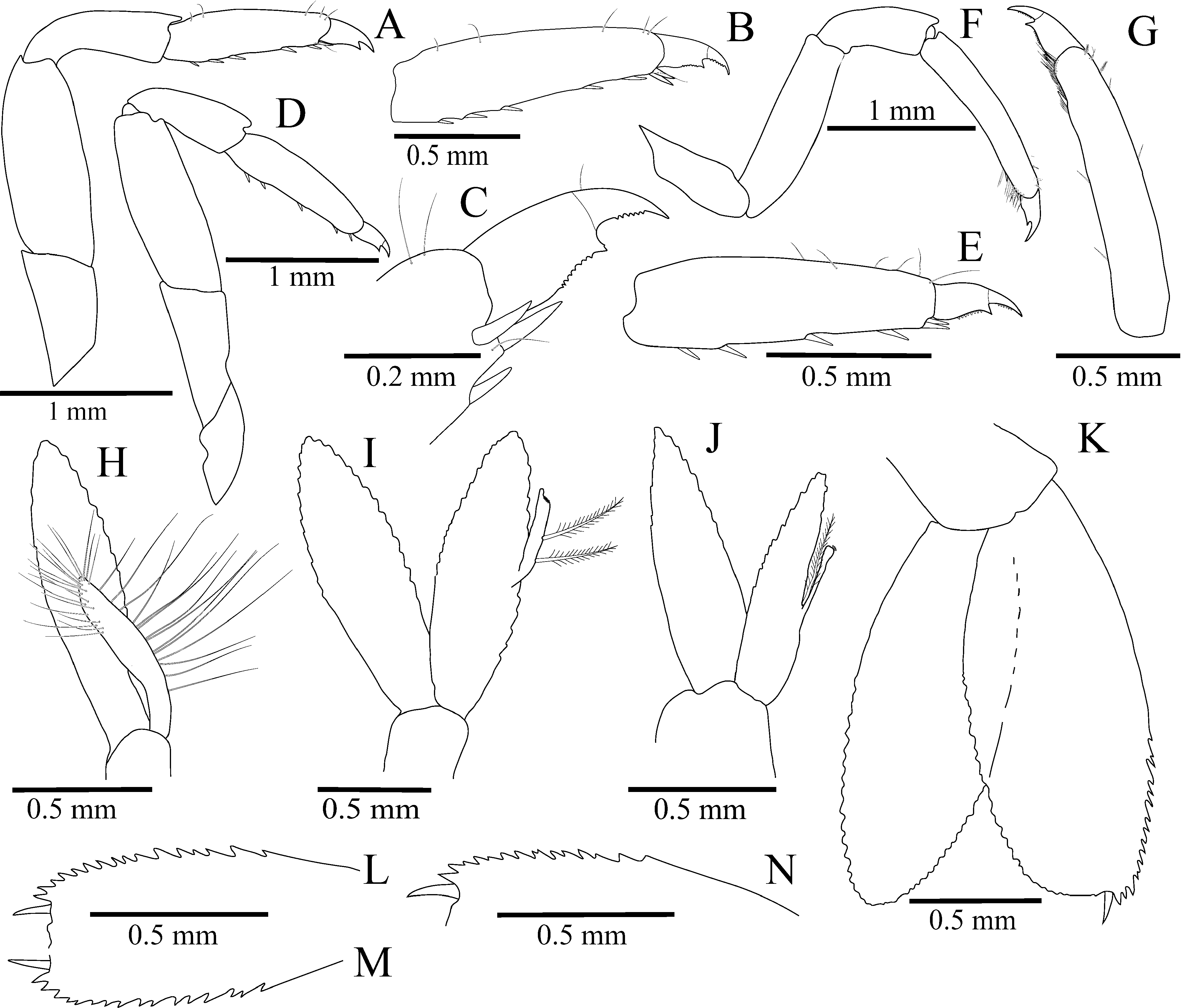
Third pereiopod ( Fig. 3 View FIGURE 3 A–C) stout; dactylus biunguiculate, compressed, 0.22 propodus length and 1.4 times longer than basal width, ventral margin of corpus slightly concave, distal half bearing minute sharp denticles and acute, triangular, ventrally pointing secondary unguis; terminal unguis distinctly demarcated, about half as long as corpus, strongly curved, bearing minute sharp denticles; propodus 3.7 times as long as wide, with pair of stout distoventral spines and 4 or 5 irregularly spaced spines along ventral margin; carpus 0.95 length of propodus, 3.2 times as long as distal width, unarmed; merus about 2.3 as long as broad and 1.2 times length of carpus, unarmed; ischium about 0.7 times length of merus, unarmed.
Fourth pereiopod ( Fig. 3 View FIGURE 3 D–E) generally similar to third pereiopod; dactylus biunguiculate, 2.5 times as long as basal width; ventral margin of corpus bearing several minute denticles in its distal half, with triangular, ventrally pointing secondary unguis; terminal unguis equal or slightly shorter than half-length of corpus, ventral margin with several minute denticles; propodus about 4 times as long as wide; ventral margin with three or 4 spines and distal pair of stouter spines; carpus 0.84 times propodus length, 2.5 times as long as distal width; merus about 2.5 times as long as wide.
Fifth pereiopod ( Fig. 3 View FIGURE 3 F–G) noticeably slenderer than third and fourth, with dactylus biunguiculate, compressed, about 0.2 of propodus length, 2.2 times as long as basal width; ventral margin of corpus slightly convex, distal half bearing minute denticles and acute, ventrally pointing secondary unguis; terminal unguis well-demarcated, slender- er than that of third and fourth pereiopods, 0.4 length of corpus, curved, with minute denticles; propodus 5.9 times as long as broad; ventral margin with 1 subdistal and 1 pair distal spines; distolateral surface with brush of grooming setae; carpus 0.6 times length to propodus, about 2.5 times as long as distal width, unarmed; merus 3.4 times as long as wide and 1.7 length of carpus, unarmed; ischium 0.7 length of merus, unarmed.
First female pleopod ( Fig. 3H View FIGURE 3 ) with endopod reaching half-length of exopod, margins with long plumose setae. Second female pleopod (Fig. 4I) about as long as exopod; appendix interna inserted at mid-length of mesial margin, with 2 long, lateral, plumose setae and about 12 subdistal cincinnuli distributed in two rows. Second male pleopod (Fig. 4J) with endopod 0.9 length of exopod, appendix interna inserted at 0.4 of mesial margin, with four subdistal cincinnuli; appendix masculina with corpus obsolete, reduced to single stiff setulose seta protruding beyond appendix interna.
Uropods (Fig. 4K–N) with protopod unarmed; exopod broadly oval, with lateral margin slightly convex, nonsetose; distolateral margin serrated in its distal third, with 14–15 acute or subacute teeth; distolateral angle with acute tooth and larger, movable spine mesially; diaeresis inconspicuous; endopod elongate, oval, slightly longer than exopod; proximal parts of lateral and mesial margins non-setose.
Color pattern. Largely transparent with yellowish-brown hue; eggs olive green.
Etymology. It is a great pleasure for the author to dedicate this species to Dr. Ana Ramos of the Spanish Institute of Oceanography, Vigo, and leader of the ECOAFRIK research team, in appreciation and recognition of her dedication to improve the knowledge of the benthic fauna of African coasts and for her inestimable support throughout the years.
Distribution. Presently known only from the type locality: Enseada de Corais da Matiota, Porto Grande Bay, São Vicente Island, Cabo Verde.
Biology. Typton anaramosae sp. nov. is, like all other species in the genus, an obligate associate of demosponges; all five specimens of the type series were extracted from unidentified sponges of yellowish-brown color and very elastic texture, probably belonging to the genus Spongionella Bowerbank.
The irritated shrimps produced a snapping sound by rapidly closing the dactylus of the major second pereiopod (major chela). The pollex of this appendage has a deep groove accommodating the thickened, almost bulging cutting edge of the dactylus. Typton spongicola and T. fapespae Almeida, Anker & Mantelatto, 2014 , both having a very similar configuration of the major second pereiopod fingers, are also known to produce snapping sounds ( Almeida et al. 2014).
Remarks. Only six species of Typton are known to have the distal part of the outer margin of the uropodal exopod serrated, viz T. spongicola (eastern Atlantic), T. holthuisi (central Atlantic), T. fapespae and T. prionurus (western Atlantic), T. serratus and T. granulosus (eastern Pacific). Typton anaramosae sp. nov. doesn’t seem to be closely related to T. spongicola , T. holthuisi and T. fapespae for it can be readily distinguished from them by the absence of a median tooth on the posterodorsal margin on the sixth pleonite. Furthermore, the serration of the uropodal exopod is restricted to a relatively small area in these three species, whilst it is over a much larger area in the new species; in addition, the number of teeth is also higher in the new species compared to the other three (cf. Bruce 2009; De Grave 2010; Almeida et al. 2014).
Typton anaramosae sp. nov. is morphologically more similar to T. granulosus , T. prionurus and T. serratus , forming a distinct species group within the genus, characterized by the outer margin of the uropodal exopod serrated in at least its distal third and the posterodorsal margin of the sixth pleonite without median tooth. Whether this species group is monophyletic remains to be tested by a phylogenetic analysis.
Typton anaramosae sp. nov. can be distinguished from T. prionurus by (1) the antennal peduncle reaching the distal margin of the second antennular article instead of reaching the end of the antennular peduncle in the latter species (cf. Fig. 1C View FIGURE 1 ; Holthuis 1951b, pl. 52, fig. a; Pachelle et al. 2015, fig. 1a–b); (2) the mesial surface of the major chela palm bearing several minute granules, which are absent in T. prionurus (cf. Fig. 2C View FIGURE 2 ; Pachelle et al. 2015, fig. 1h); (3) the much higher number of teeth in the serrated margin of the uropodal exopod (cf. Fig. 3 View FIGURE 3 K–N; Holthuis 1951b, pl. 52, fig. c; Pachelle et al. 2015, fig. 1m); (4) the slenderer first pereiopod, exceeding the antennular peduncle by the distal third of the merus, instead of by the length of the carpus in T. prionurus (cf. Fig. 1I View FIGURE 1 ; Holthuis 1951b: pl. 52, fig. h); (5) the relatively shorter first pereiopod fingers, with finger-palm ratio equal to 0.45 in the new species versus fingers slightly more than half length of the palm in T. prionurus ; (6) the dactylus of second major cheliped proportionally shorter in comparison to the palm length (0.34–0.37 vs. 0.5); (7) the scaphocerite terminally rounded and reaching to about 0.3 length of first article of the antennal peduncle vs. pointed and reaching distal third of first article of the antennal peduncle in T. prionurus (cf. Fig. 1G View FIGURE 1 ; Holthuis 1951b, pl. 52, fig. e); (8) the carpus of second major cheliped slightly more produced ventrally, being 1.3–1.4 times as long as maximal width vs. about 1.7 times as long as maximal width in T. prionurus ; (9) the dactylar corpus of the third pereiopod with denticles restricted to its distal half vs. extending for almost its entire length in T. prionurus (cf. Fig. 3C View FIGURE 3 ; Pachelle et al. 2015, fig. 1l); (10) the third pereiopod dactylus stouter; and (11) the third pereiopod carpus somewhat larger in comparison to the propodus (0.95 vs. 0.83) and merus (0.84 vs. 0.71) (cf. Fig. 3A View FIGURE 3 ; Holthuis 1951b: pl. 52, fig. k).
Typton anaramosae sp. nov. can be distinguished from T. granulosus by (1) the slenderer first pereiopod, with the carpus being about 6.6 and the merus 7.4 times longer than wide vs. 4.7 and 6.4 times longer than wide, respectively, in T. granulosus ; (2) the much higher number of teeth on the serrated margin of the uropodal exopod (cf. Fig. 3 View FIGURE 3 K–N; Ayón-Parente et al. 2015, fig. 5D–I); (3) the presence of several minute denticles on the unguis of the third to fifth pereiopod dactylus (cf. Fig. 3 View FIGURE 3 B–C, E, G; Ayón-Parente et al. 2015, fig. A–F); (4) the relatively length of the scaphocerite, reaching to about 0.3 length of first article of the antennal peduncle vs. reaching to its half-length in T. granulosus (cf. Fig. 1G View FIGURE 1 ; Ayón-Parente et al. 2015, Fig. 2B View FIGURE 2 ); (5) the somewhat broader telson, about twice as long as wide in the new species vs. 2.5 times longer than wide in T. granulosus ; and (6) the smaller spines on the dorsal surface of the telson (0.08 of telson length in T. anaramosae sp. nov. vs. 0.14 in T. granulosus ) (cf. Fig. 1E View FIGURE 1 ; Ayón- Parente et al. 2015, fig. 1G).
Finally, T. anaramosae nov. sp. can be easily distinguished from T. serratus , for instance, by (1) the second pair of dorsal spines of the telson set in its posterior half vs. both pairs closely set together in the anterior quarter of the telson in T. serratus (cf. Fig. 1E View FIGURE 1 ; Holthuis 1951b: pl. 53, fig. c); (2) the slenderer first pereiopod, exceeding antennular peduncle by the distal third of the merus vs. exceeding that article by only part of carpus in T. serratus ; (3) the major cheliped palm slightly convex on dorsal and ventral margins vs. with straight margins in T. serratus (cf. Fig. 2 View FIGURE 2 A–C; Holthuis 1951b: pl. 53, fig. i); (4) the uropodal exopod serration with more smaller teeth and starting at about 0.6 length of the outer (lateral) margin vs. with fewer and larger teeth, starting at mid-length of the outer margin in T. serratus (the uropodal exopod also being broader and more ovate in the new species) (cf. Fig. 3 View FIGURE 3 K–N; Holthuis 1951b: pl. 53, fig. c); and (5) the noticeably shorter rostrum (cf. Fig. 1 View FIGURE 1 A–B; Holthuis 1951b, pl. 53, fig. b).
Acknowledgments
I am indebted to Sandra Correia and Marcia Costa (Instituto do Mar—former Instituto Nacional para o Desenvolvimento das Pescas, Mindelo, Cabo Verde) for their help and arrangements during the use of facilities and equipment during the description of the new species. I am also grateful to Paulo Pachelle, Sammy De Grave and Zdeněk Ďuriš for corrections and very useful comments during the preparation of the manuscript and (S. De Grave) for correction of the English language. I am deeply grateful with the comments and suggestions of A. Anker and other anonymous reviewer that help to improve the quality of this manuscript
| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |