
Circunula, Koppka, Jens, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3927.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:42B56D11-9B18-4FCC-B632-30A46AB0205C |
|
DOI |
https://doi.org/10.5281/zenodo.6102690 |
|
persistent identifier |
https://treatment.plazi.org/id/E502D251-D402-446E-A9AB-E60888911559 |
|
taxon LSID |
lsid:zoobank.org:act:E502D251-D402-446E-A9AB-E60888911559 |
|
treatment provided by |
Plazi |
|
scientific name |
Circunula |
| status |
gen. nov. |
Circunula n. gen. cotyledon ( Contejean, 1859)
Figs. 5–8 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 ; Pls. 1–2; Pl. 3.1–5; Pl. 4.1, 4
? 1837 Ostrea multiformis , nobis. var. orbicularis Koch & Dunker : p. 45, pl. 5, fig. 11n (only). v * 1859 Ostrea cotyledon, Ctj. Contejean : p. 319, pl. 24, fig. 15–17. v 1861 Ostrea cotyledon, Ctj. —Thurmann & Etallon: p. 45, 53, 55, 56, 61, 66.
1862 Ostrea cotyledon, Ctj. — Etallon: p. 442.
1862 Ostrea cotyledon, Ctj. —Thurmann & Etallon: p. 271, pl. 39, fig. 2.
1867 Ostrea cotyledon, Ctj. —Greppin, J.-B.: p. 92.
1867 Ostrea cotyledon, Ctj. —Moesch: p. 200.
1870 Ostrea cotyledon, Ctj. —Greppin, J.-B.: p. 82, 104.
1872 Ostrea cotyledon Contejean—de Loriol, Royer & Tombeck: p. 406, pl. 24, fig. 2.
1880 Ostrea cotyledon Contej. —Struckmann: p. 62.
1882 Ostrea multiformis Dunk. et Koch—Alth : p. 296 (114), pl. 27 (10), fig. 20.
1890 Ostrea cotyledon, Ctj. —Mathey: p. 17.
1905 Ostrea cotyledon Contejean—Schmidt : p. 159.
1917 Ostrea cotyledon Contej. —Rollier: p. 590.
1963 Liostrea cotyledon Contej. —Pčelinčev & Lysenko: p. 133.
1971 Liostrea cotyledon —Pugaczewska: p. 198.
v 1973 Ostrea cotyledon Ctj. —Contini & Hantzpergue: p. 175. v 2006 Liostrea cotyledon—Hicks: p. 36.
v 2006 Liostrea cotyledon ( Contejean 1859) —Richardt: p. 9, 18.
Lectotype (designated herein). Ostrea cotyledon Contejean, 1859 (pl. 24, figs. 15–16), MC-27-E-105: Bivalved specimen growing on a fragment of Trichites sp., with attached younger LV (paralectotype) MC-27-E-105a (internal features uncovered by present author) (Fig. 5.1–3). Rôce in Sainte-Suzanne, Montbéliard (Département Doubs, Franche-Comté), France. Coordinates ca. 47°30'20.33''N, 6°47'51.00''E. The exact position of the locality is unknown.
Lower Kimmeridgian, cymodoce Zone; “Niveau 6” of Contejean (1859), "Calcaires et Marnes à Ptérocères” (= "Marnes du Rang" in eastern France), equivalent of the Banné Member of the Swiss Reuchenette Formation, Upper Jurassic.
Material. Lectotype (see above, MC 27E-105), paralectotypes MC 27E-105a and MC 27E-104 (from type locality) (Fig. 5.1–5). Paralectotype MJ 27E-108 from Arbouans (between Montbéliard and Audincourt), Upper Oxfordian “ Natica Limestone ” ("Calcaires à Natices", niveau 2 of Contejean (1859), equivalent of the Vorbourg Member, Lower Vellerat Formation of Switzerland); Musée Cuvier in Montbéliard (Fig. 5.7–9).
Others—Numerous specimens from the Banné Marls (cymodoce Zone) from Vâ Tche Tchâ ( VTT), Alombre aux Vaches (ALO), Cras d’Hermont (CRA), Chevenez-La Scierie (CHS) in northwestern Switzerland and several specimens from a hardground at the base of the lallierianum Subzone (mutabilis Zone) of Sur Combe Ronde (SCR). One specimen from Blauen (Upper Oxfordian) of the Thurmann Collection (S 1469, = A.1.10.5.7; Pl. 3.19) in the MJSN Porrentruy (Appendix, Table 1).
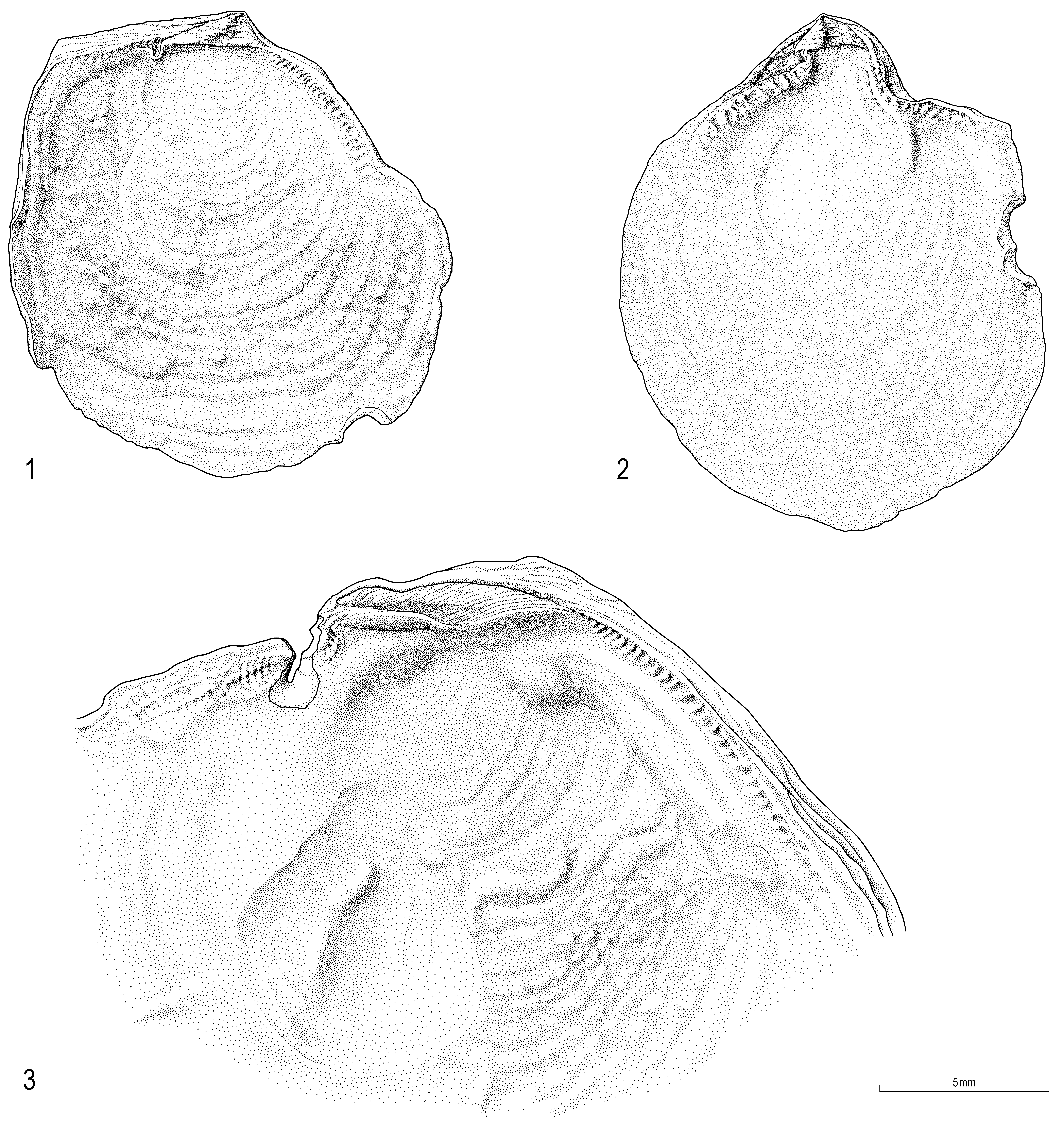
Description. Shape, size—Left valve almost entirely cemented, larger than RV; ventral shell margin may become detached from surface, bending upwards at angles between 90° and 110°; posteroventral margin usually highest (see paleoecology); RV rather flat, ventrally partly concave, commissural shelf bending upwards, emulating free LV margin (Pl. 1.5d, Pl. 4.1, 4). Examined specimens between 1.1–7.5 cm high (Appendix, Table 1). Sculpture—Free part of LV with fine, even-spaced commarginal growth steps (Pl. 1.3b, 5c); very thin shells occasionally revealing external sculpture of attached surface consisting of minute, discontinuous, partly bifurcating, antimarginal and concentric furrows, few antimarginal riblets, and pustules (Pl. 2.5a,b,e, 7a,b) (Pl. 3.3b,c,e).
RV with weak commarginal growth lines and steps, and few antimarginal riblets (only on well preserved shells) (Pl. 2.2b, 4b, 5b, 6), becoming weakly squamous towards ventral margin (Pl. 1.5b); xenomorphic sculpture rare (smooth attachment surfaces preferred) (Pl. 1.5a,b).
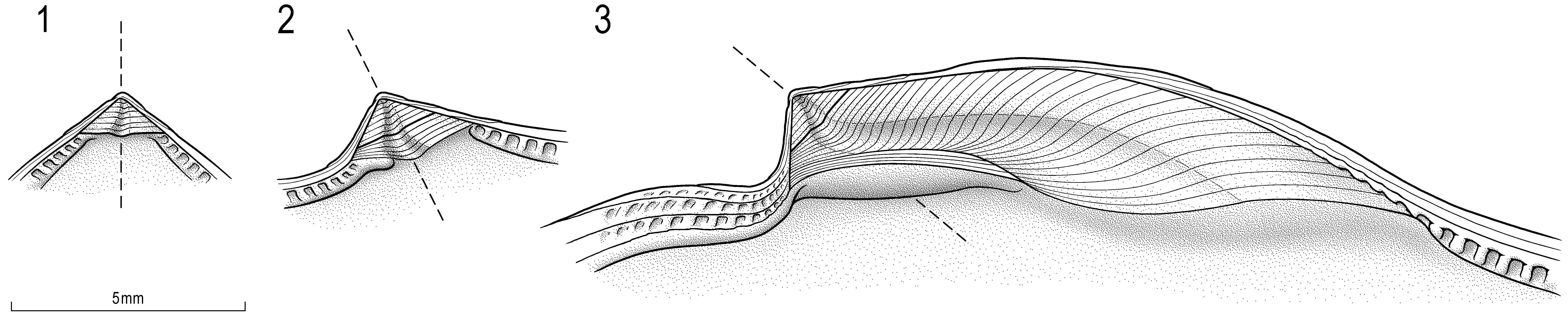
Ligament area—Small, more or less triangular, alivincular-areate (Fig. 6.1–2); some shells with an initial prosogyrate twist (Fig. 6.1; Pl. 3.3b, 4–5), (Fig. 7.1–3; Pl. 3.4); resilifer weakly concave with slightly convex bourrelets in both valves, but less elevated in RV; early growth stages overgrown by projection of posterodorsal hinge margin (Fig. 7.2–3).
Internal shell characters—Umbonal cavity lacking. Posterior adductor scar (PAM) round, except dorsally, oblate (Pl. 1.1b) to concave (Pl. 2.1); position in adults posterodorsal, close to shell centre. Large specimens occasionally with deep sickle-shaped gill depression between posterior shell margin and adductor scar, reaching from hinge to branchitellum (LV: Fig. 5.9; Pl. 2.1; RV: Fig. 5.7; Pl. 1.1b; Pl. 3.1).
Microstructure—RV with thin (ca. 170 µm) continuous outer shell layer of reclined prisms; middle-inner shell layers low-angle foliate without chambering. Prisms and sculptures, commonly bioeroded through grazing by regular echinoids (star-shaped trace fossil Gnathichnus pentax Bromley, 1975 ; Pl. 2.4b). Left valve not examined but apparently lacking chambers or vesicular shell layers (no indications under light microscope).
Prodissoconch, juvenile—Prodissoconch and earliest postlarval shell not preserved. “Juveniles” or smallest adults typically with strongly gyrate ligament area, usually opisthogyrate, but prosogyrate in 7 out of 22 specimens. Posterior bourrelet may be very narrow (Fig. 6.3). Shape of juvenile PAM often more crescentic to subrectangular than round and slightly closer to hinge than in adult ( Fig. 7 View FIGURE 7 ; Pl. 3.3b,c, 5). Small linear chomata on both sides of dorsal margin ( Figs. 6–7 View FIGURE 6 View FIGURE 7 ; Pl. 3.2–5) down to height of adductor scar; anterior catachomata (LV) as parallel grooves (length 0.17–0.3 mm, width 0.09–0.17 mm), normal to and along commissural shelf. Posterior catachomata present, larger specimens with relict catachomata especially close to hinge but not always well developed. Chomata overgrown and absent in larger shells (Fig. 7.3; Pl. 1.1b, 4b). Most juveniles and small adults smooth internally ( Figs. 6 View FIGURE 6 , 8 View FIGURE 8 ; Pl. 3.3a, 4–5).
Remarks. Coiling—Early postlarval prosogyry is relatively common in Circunula cotyledon (e.g. VTT001- 3215, VTT001-3238, VTT001-1613); this tendency is perhaps only matched by Jurassic Catinula knorri ( Voltz, 1828) (Pl. 4.2, 3b). Prosogyry appears to be somewhat more common among specimens settling on the dark (protected?) interior of a bivalved Trichites shell (ALO009-1) or on the underside of Isognomon specimens (VTT001-1615, VTT001-3215), but overall observations do not support an environmental influence.
Chomata— Circunula cotyledon is the first Jurassic species with chomata that does not belong to Exogyrinae , Arctostreidae , or to Pseudeligmus ( Malchus 1990, 1998, p. 402, fig. 5). Their restriction to the early postlarval growth phase and later overgrowth and disappearance is not unusual, however.
Paleoecology. Circunula n. gen. cotyledon is a typical pioneer settler on limestone hardgrounds such as the basal hardground of the Banné Member, and layers 2000, 3000 of the Sur Combe Ronde section ( Fig. 4 View FIGURE 4 B–C) where they become largest, develop the thickest shells and usually remain entirely attached (Pl. 1.2, Pl. 2.1, 7). They also occur commonly on Isognomon shells (Pl. 2.3) in oolitic and biomicritic sediments of the basal Banné Marls. Optimal conditions thus appear to have been medium to high energy environments in shallow marine carbonate settings, where most of the other oyster species could not survive.
Small articulated adults (2–3 cm) with a raised ventral margin are more typical of the highly fossiliferous marls of the middle Banné Member (Pl. 1.3, Pl. 2.4, Pl. 4.1), settling on large, smooth shell surfaces of Isognomon , Ceratomya (Pl. 1.5), and Trichites (Pl. 3.5). These deposits likely indicate calmer conditions.
Figure 8 View FIGURE 8 and Plate 3.2–3 show a larger slab of a Trichites shell (ALO009-1, bivalved specimen) with post mortem settlement by more than 20 juvenile specimens. Overall, shells are attached with their ventral side directed towards the ventral margin of the Trichites shell. Specimens are much larger close to the margin than those attached more dorsally ( Fig. 8 View FIGURE 8 ). This orientation may indicate preferential growth close to nutrient-rich incoming current. The idea is supported by similarly oriented specimens of Nanogyra (N.) nana attached on the outer surface of the same Trichites shell (Pl. 6.1a – c).
Occurrence. Upper Oxfordian to Upper Kimmeridgian of the Reuchenette Formation in Switzerland; Upper Oxfordian–Kimmeridgian in France; Kimmeridgian in Germany, Poland, Russia (Crimea); Tithonian in the Czech Republic.
Comparisons. The generic assignment of species discussed below remains tentative. The first three species could belong to Circunula , but there is no evidence of chomata, so far. Several more Liostrea -like species are discussed afterwards in alphabetical order.
Liostrea (Catinula) stoliczkai Cox, 1952 (p. 75, pl. 6, figs. 5–6)—This species was originally described by Cox (1952) from the Upper Bathonian of Soorkha (Kutch district, Gujarat, India). Morphological similarity with C. cotyledon may indicate that the two species are congeneric.
The species differs by having a tendency to develop a more quadrate outline , by being more strongly reflected upwards and growing higher ventrally (up to ca. 4 cm; fide Cox 1952, p. 75). The RV shows better developed antimarginal riblets, occasionally also radial furrows which reach near the ventral margin, and commarginally arranged tiny pustules ( Cox 1952, pl. 6, fig. 6a). Remains of such pustules in C. cotyledon are only known from juvenile LVs (Fig. 6.1, 3; Pl. 3.3b,c,e). Judging from the figured LV in Cox (1952, pl. 6, fig. 6b), the PAM appears to be larger and more centrally situated than in C. cotyledon .
" Liostrea " strigilecula ( White, 1877) (p. 163, pl. 8, fig. 3a–d)—This species was described from the Middle Jurassic Carmel Formation in SW Utah. It has a much larger adductor and a more inflated LV than C. cotyledon . It produces free-rolling oyster accumulations (ostreoliths) in shallow marine environments and also grows on hardgrounds ( Wilson et al. 1998, p. 73, fig. 4A).
“ Ostrea ” planaria Rollier, 1917 (p. 550, pl. 39, figs. 1a–c)—The species comes from the "Terrain à Chailles" (Lower Oxfordian, cordatus Zone) of the ancient "Bisthum Basel" ( Switzerland) and the environment of Ferrette (Pfirt) in the Alsace, France.
The specimen figured in Rollier (1917) has a smaller attachment area and the ventral margin is less towering than in C. cotyledon . Both sides of the complete specimen are encrusted by serpulids and several specimens of Nanogyra (probably N. nana ) suggesting a life mode under low energy conditions. Small, poorly preserved Circunula -like shells found by the author attached to corals in the somewhat younger marls and limestones of the Liesberg Member (N Switzerland) probably belong to the same species. It may be congeneric with C. cotyledon .
“ Ostrea ” matisconensis Lissajous, 1923 (p. 123, pl. 28, figs. 9–12)—This is a Bathonian species from France. Similar to C. cotyledon , it has a circular outline but it remains much smaller and develops antimarginal riblets on the unattached part of the ventral LV margin. These riblets may indicate an assignment to Catinula View in CoL rather than Circunula .
" Ostrea " matronensis de Loriol in de Loriol, Tombeck & Royer, 1872 (p. 396, pl. 23, figs. 5–7)—The species occurs in the Portlandian ( gigas Zone, Tithonian ) of the Department Haute-Marne, France. It is characterized by a less oblique opisthogyrate resilifer and a higher ligament area; the umbo is more prominent and the overall shape tends to be more subtriangular than round. De Loriol (1872, pl. 23, figs. 6, 6a) figured a specimen attached to an ammonite, but otherwise settling grounds are unknown.
? Liostrea moreana ( Buvignier, 1852) (p. 26, pl. 16, figs. 41–43)—This large species (up to 13 cm) was described from the Oxfordian (“coral rag”) of the Meuse Department (Lorraine) in northeastern France. Differences to C. cotyledon are the larger adult size, the more capacious and stronger upward reflected left valve (up to 8 cm), a broader ligament area with a ventrally straight RV hinge margin, and absence of antimarginal ornament on the RV.
“ Ostrea ” sandalina Goldfuss, 1833 (b) (pl. 79, figs. 9a–m)—The material from Goldfuss appears to belong to various species. Specimens of figures 9c–d come from a “black Jura-limestone” (Lower? Jurassic) of Osterkappeln and Lübke, NW Germany; specimens 9a–b, f–I are from oolitic ferruginous clay stones (probably Bajocian) of Streitberg, Gräfenberg, and Thurnau, Bavaria. Goldfuss also mentioned this species from white Oolites (Upper Jurassic) of Hildesheim and Goslar, northern Germany.
His specimens of figures 9c–d, h, l–m may indeed belong to Circunula n. gen. whereas figures 9i –k more likely represent Liostrea or Praeexogyra . Compare also specimens tentatively determined as? Praeexogyra sandalinoides (de Loriol, 1901) from the Callovian of Western Pomerania (Pl. 15.3–4).
Cox (1952, p. 73, pl. 6, figs. 1–4, with synonymies) described material from the Bathonian of India and determined the species as Liostrea (Catinula) sandalina . However, typical specimens with a large attachment area and oval outline differ from C. cotyledon by a smaller adult height (around 3 cm), a less orbicular shape and a somewhat stronger upward reflected ventral margin.
Liostrea brasili Chavan, 1952 (p. 43, pl. 2, fig. 22)—The species from the Oxfordian of Calvados ( France) has a similar shape, but a triangular ostreoid hinge, with a broad resilifer and a ventrally convex hinge line. It also has a well-developed umbonal cavity ( Chavan 1952, pl. 2, fig. 22), which is absent in Circunula n. gen.



| VTT |
VTT Biotechnology, Culture Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Circunula
| Koppka, Jens 2015 |
Liostrea (Catinula) stoliczkai
| Cox 1952 |
Liostrea brasili
| Chavan 1952 |
Liostrea moreana (
| Buvignier 1852 |
sandalina
| Goldfuss 1833 |