
Trachelostenus Solier, 1851
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4020.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:7FBBE9CA-E98F-4638-8A5E-7E51816B6B79 |
|
DOI |
https://doi.org/10.5281/zenodo.6097778 |
|
persistent identifier |
https://treatment.plazi.org/id/4F43853E-841C-FFFB-FF5D-238445D5D4C8 |
|
treatment provided by |
Plazi |
|
scientific name |
Trachelostenus Solier, 1851 |
| status |
|
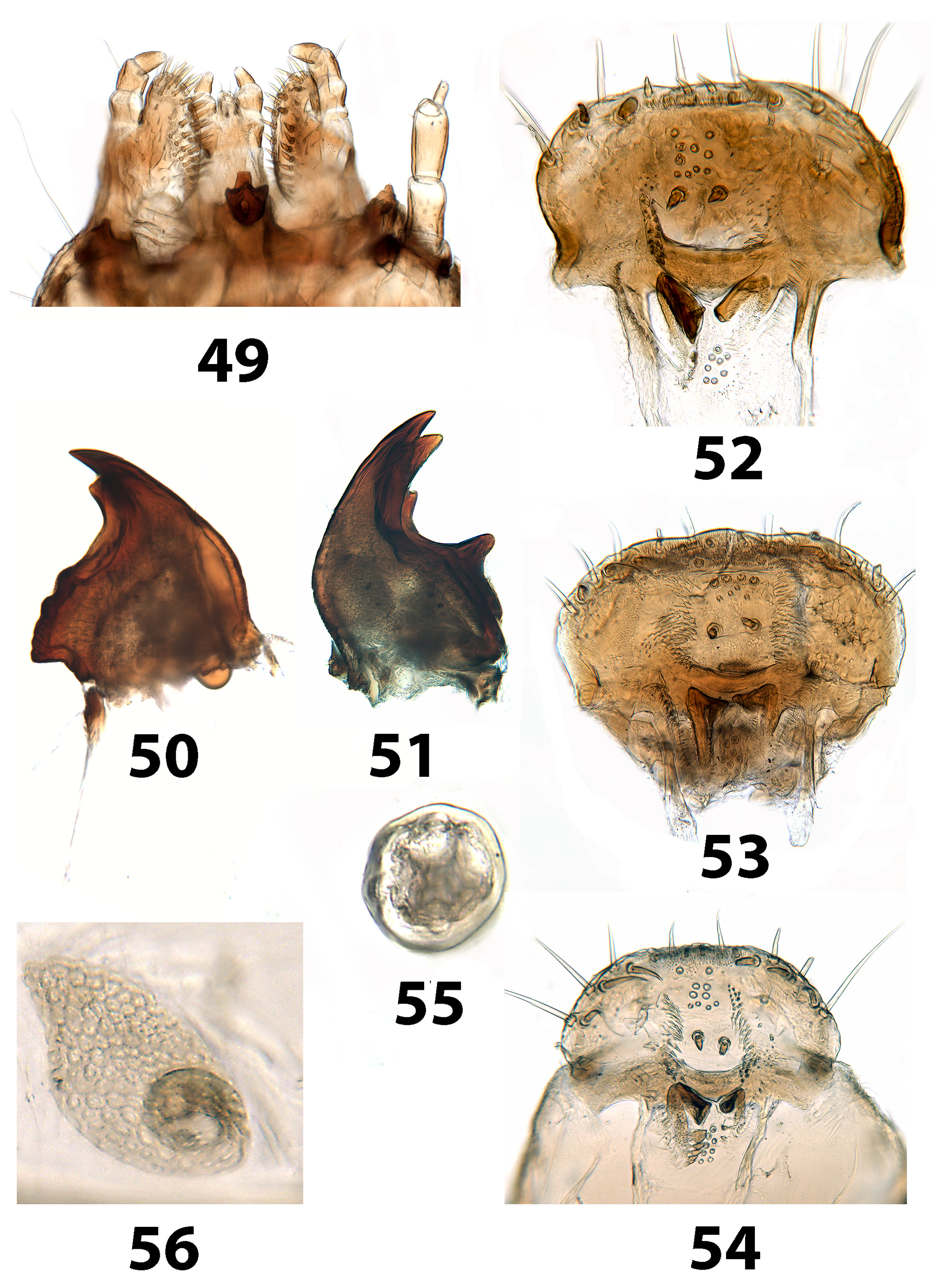
( Figs 1 View FIGURES 1, 2 , 3 View FIGURES 3 – 5 , 6 View FIGURES 6, 7 , 8 View FIGURES 8 – 10 , 11 View FIGURE 11 , 12–16 View FIGURES 12 – 14 View FIGURES 15, 16 , 38, 41 View FIGURES 37 – 41 , 42, 45 View FIGURES 42 – 48 , 49, 52 View FIGURES 49 – 56 )
Type species: Trachelostenus inaequalis Solier, 1851 by monotypy.
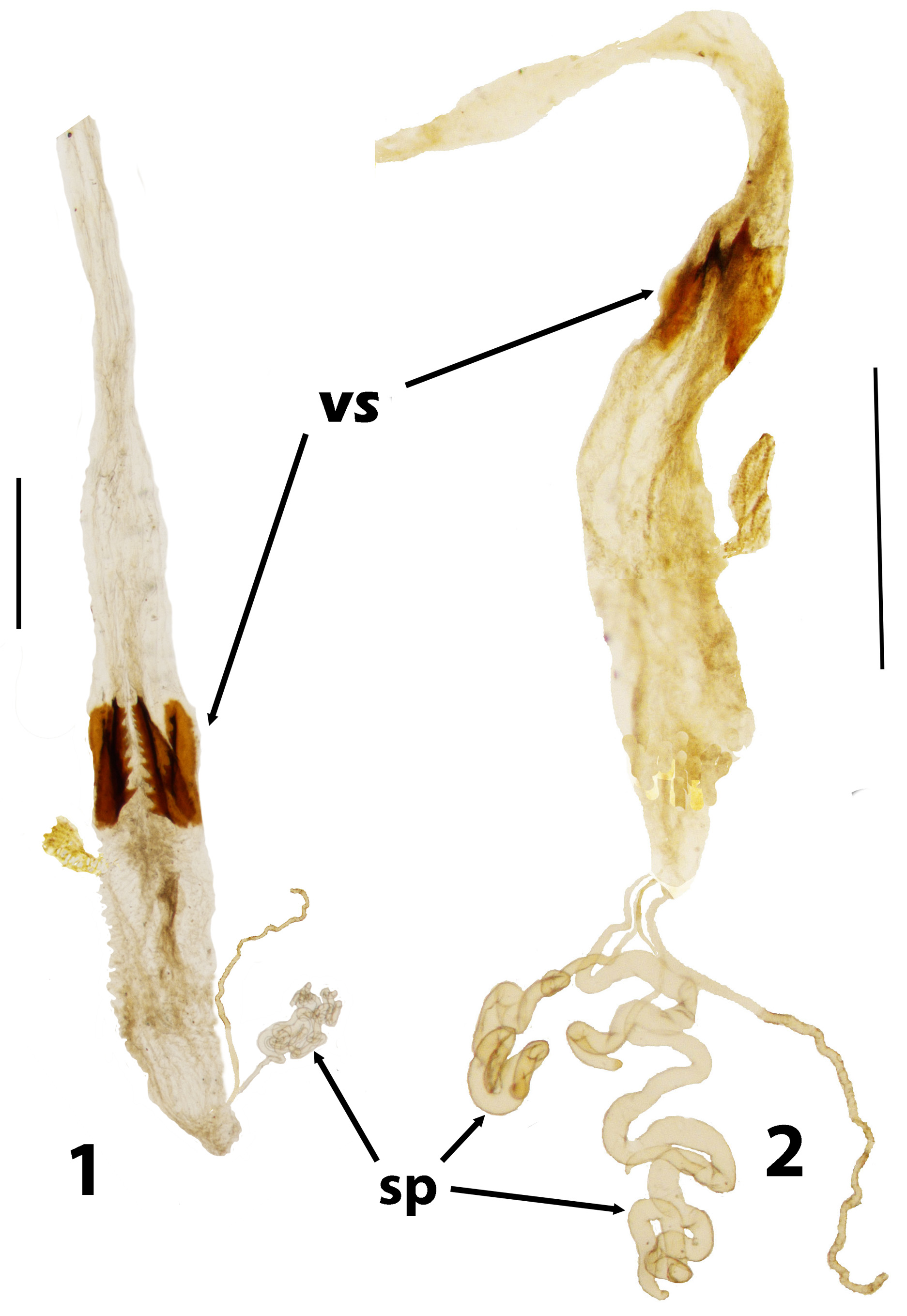
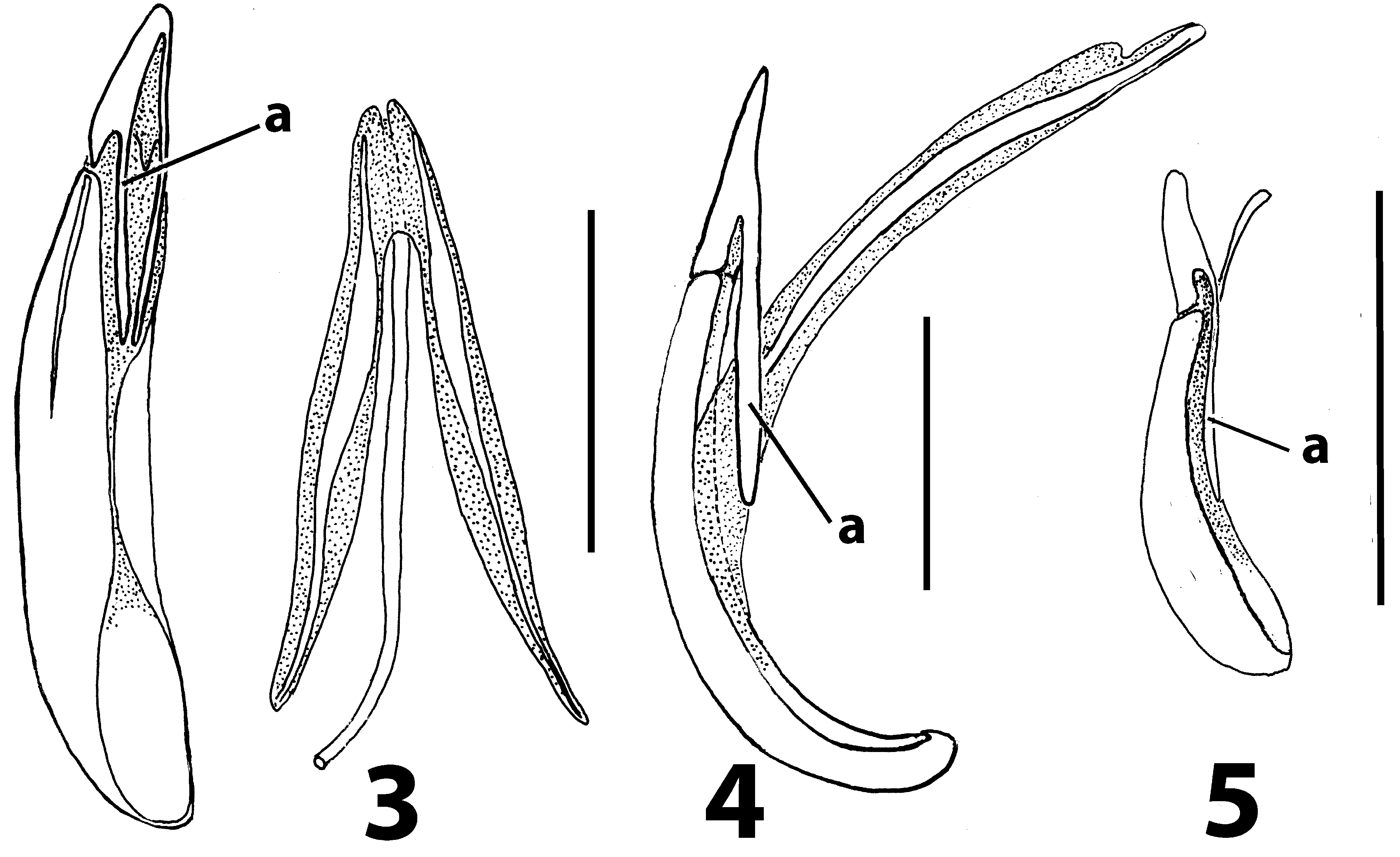
Redescription. Adults. With the characteristics of the tribe. Length 6.5–10.5 mm. Body parallel-sided, 3.3– 3.6 times as long as wide, with prothorax narrower than elytral bases, fuscous, legs with alternating flavous and fuscous bands. Upper surfaces glabrous or clothed with patches of semierect white hairs, or sparse semirecumbent pilosity. Head: Basal membrane of labrum moderately wide. Eyes shallowly emarginate anteriorly. Frons with deep median pit. Antennae very long, filiform, reaching to about anterior third of elytral length, antennomeres elongate, terminal one oblong, not enlarged. Mentum transverse, about twice as wide as long. Prothorax: Pronotum oblong, sides subparallel, almost straight or a little convex, lateral carinae absent. Procoxal cavities internally closed by very slender bar. Concealed propleurotrochantins present. Pterothorax: Elytral disc with elevations, at least anteriorly. Wings fully developed. Legs: Pro- and mesocoxae subconical and prominent. Procoxae projecting well below prosternum, approximated so as to cover latter. Mesocoxae prominent and narrowly separated. Femora and tibiae long and slender, tarsomeres linear except penultimate, which is slightly widened, together as long as or longer than tibiae. Abdomen: Intercoxal process of first ventrite narrow, subparallel or triangular, with rounded apex. With only first 2 ventrites connate. Tergite VII with a pair of finely rugose patches in both sexes. Defensive glands absent. Male: In T. inaequalis with a deep longitudinal groove joining frontal pit with fronto-clypeal suture, not present in other species. Tergite VII a little longer than wide. Sternite VIII arcuately excised, in one unnamed species with outer angles of excision prolonged into short spurs. Sternite IX partly or completely divided into 2 sclerites, Female: Tergite VII about as long as wide. Ovipositor with paraproct baculi subparallel to axis of ovipositor, baculi of first coxite lobes also parallel, about 1.5 times as long as paraproct baculi. Coxites with only 3 pairs of lobes, last 2 weakly sclerotised, rest of ovipositor membranous except for baculi. Vaginal sclerites (of T. inaequalis ) in form of 4 coarsely-toothed saw blades. Spermatheca a tightly coiled tube attached to basal part of accessory gland.
Adult material examined: T. inaequalis . CHILE: Cudico Villarrica, 27-I-1978 ( FMNH) (1); Osorno Prov. Parque nac. Puyehue, 4.1 km E Anticura, 430 m Site 662, 19-26 Dec. 1982, Valdivian RF, Newton & Thayer ( FMNH) (1); 30 km E of Puyehue, Osorno ( CAS) (1); Arauco Caramavida ( FMNH) (1); Chaitén Aysén, XII-1985 ( FMNH) (3); Cherquenco [abdomen only] ( FMNH). T. fasciculiferus . La Selva 700m Nueva Imperial, 11-81, Coll. L. E. Peña ( FMNH) (3). One unnamed species. Tepuhuelco Chiloe ( FMNH) (1).
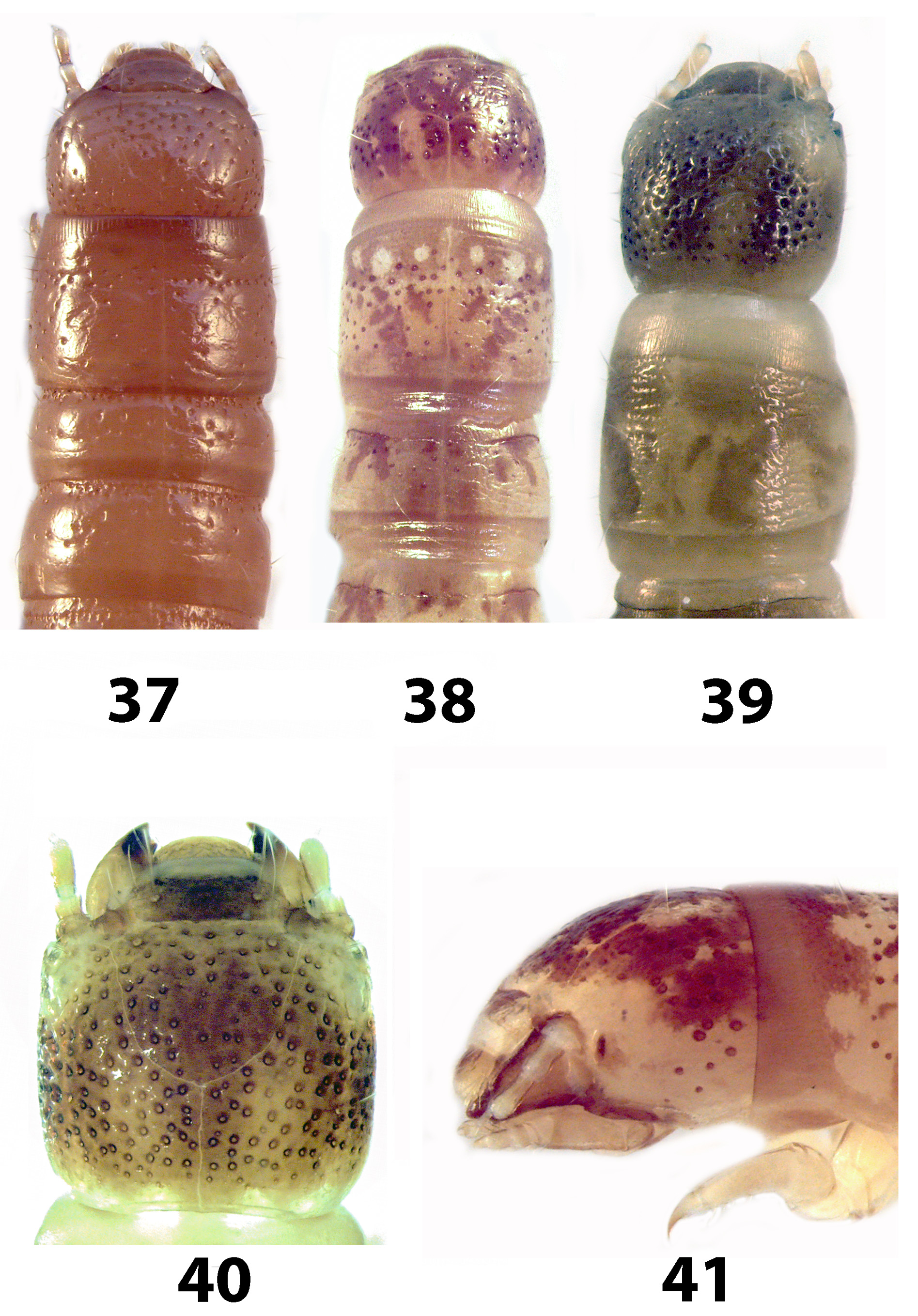
Redescription. Larvae. With the characteristics of the tribe. Dorsal surfaces bicolored with a complex pattern of dark brown markings on a yellow background. Head distinctly rounded; frontal arms somewhat irregularly curved and angulate; larger anterior epipharyngeal sensilla with anterior 2 moderately widely separated and close to the posterior 4, which form a sharply angulate row ( Fig. 52 View FIGURES 49 – 56 ); posterior sensilla forming an irregular cluster; mandibles bidentate with short subapical tooth on incisor edge; left mola with several weak transverse ridges; gula slightly shorter than wide; hypopharyngeal sclerome anteriorly trilobed. Prothorax almost as long as wide. Legs slightly longer than thoracic width and clothed with fine hairs. Abdomen about 3.5 times as long as thorax; tergum IX distinctly longer and slightly wider than VIII, subcircular carina simple ( Fig. 45 View FIGURES 42 – 48 ); urogomphi each with 4 setiferous tubercles at about middle; surface of concave disc lightly pigmented; segment X with a pair of tubular pygopods. Spiracles annular-multiforous with a variable number of accessory chambers located around the peritreme (as in Fig. 55 View FIGURES 49 – 56 ).
Larval material examined: T. inaequalis . CHILE: VIII: Parque Nac. Nahuelbuta, Pichinahuel Exit, 1219m, 08.xii.2001, fogging Nothofagus dombeyi, E. Arias et al. U.C. Berkeley Expedition ( ANIC); X: Osorno: 4.1 km E Anticura, 430m, 19–26.xii.1982, A. Newton & M. Thayer colls ( ANIC). Trachelostenus sp. IX: Parque Nac. Villarrica, Sector Puesco, 873m, 16.i.2007, under bark of logs, J. F. Lawrence coll. ( ANIC).
Distribution and habits. The above localities are in three of the biogeographic regions of Chile recognised by Vidal & Guerrero (2007): Bosque de Valdivia del Norte, Bosque de Valdivia, and Cordillera de Aysén. Nothophagus forest is the dominant vegetation type. Philippi & Philippi (1860) describe the habitat of T. fasciculiferus as ‘ in Valdivia sub cortice arborum rarissime ’ and Lawrence & Slipinski (2010) also note that adults are found under bark, but the variegated pigmentation of the larvae suggests external feeding, possibly on lichens as in Titaenini.
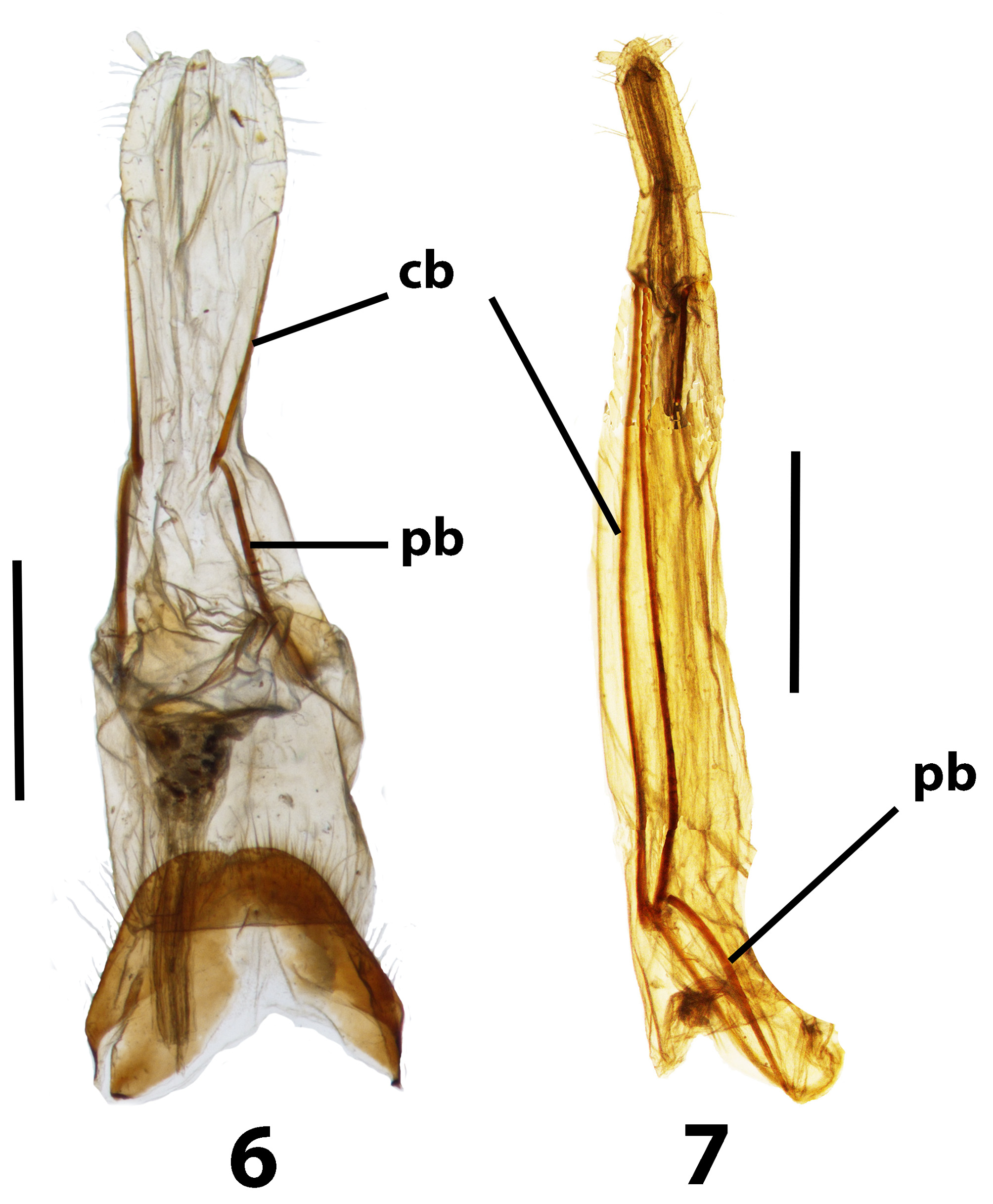
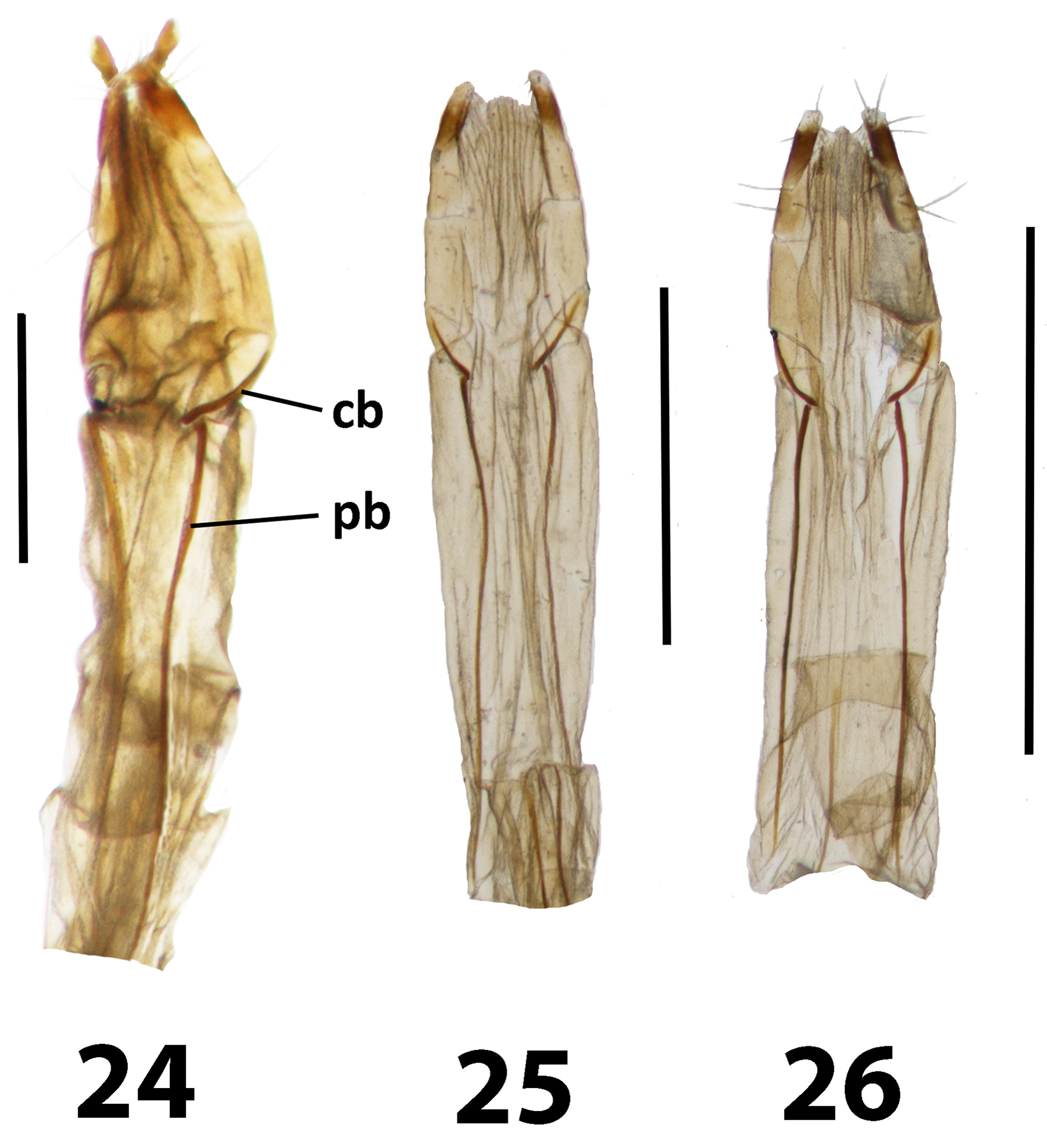
Discussion. Evidently Trachelostenus has lost the ancestral defensive glands and with them the connation of the third ventrite ( Fig. 8 View FIGURES 8 – 10 ). There is a striking apomorphy which is shared with Myrmecodema : the unique ovipositor structure with longitudinal and elongate coxite-1 baculi ( Figs 6, 7 View FIGURES 6, 7 cb), first noted by Lawrence & Slipinski (2010) in Trachelostenus . Normally in Tenebrionidae coxite-1 baculi and lobes are short and transverse or diagonal, as they are in Leaus for instance ( Figs 24–26 View FIGURES 24 – 26 cb). Otherwise, longitudinal coxite-1 baculi are seen only in the distinctive ovipositor of Stenochiinae as first described by Tschinkel & Doyen (1980). However, the stenochiine ovipositor has completely transverse paraproctal baculi, whereas these are more or less longitudinal in Trachelostenus and Myrmecodema . In addition, the stenochiine ovipositor has a pair of newly evolved, extra baculi joining the ends of the paraprocts with the proctigeral baculi. Trachelostenini do not have these extra baculi. Therefore, the particular ovipositor structure of Trachelostenus and Myrmecodema is unique among known tenebrionid ovipositors.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |