
Teleiopsis sharporum, Bidzilya, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4952.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:E32A9743-839A-4740-A8B7-E82340238097 |
|
DOI |
https://doi.org/10.5281/zenodo.4700931 |
|
persistent identifier |
https://treatment.plazi.org/id/F53D6A1C-A321-6544-5A9A-F95F6DAA5B5A |
|
treatment provided by |
Plazi |
|
scientific name |
Teleiopsis sharporum |
| status |
sp. nov. |
Teleiopsis sharporum View in CoL sp. nov.
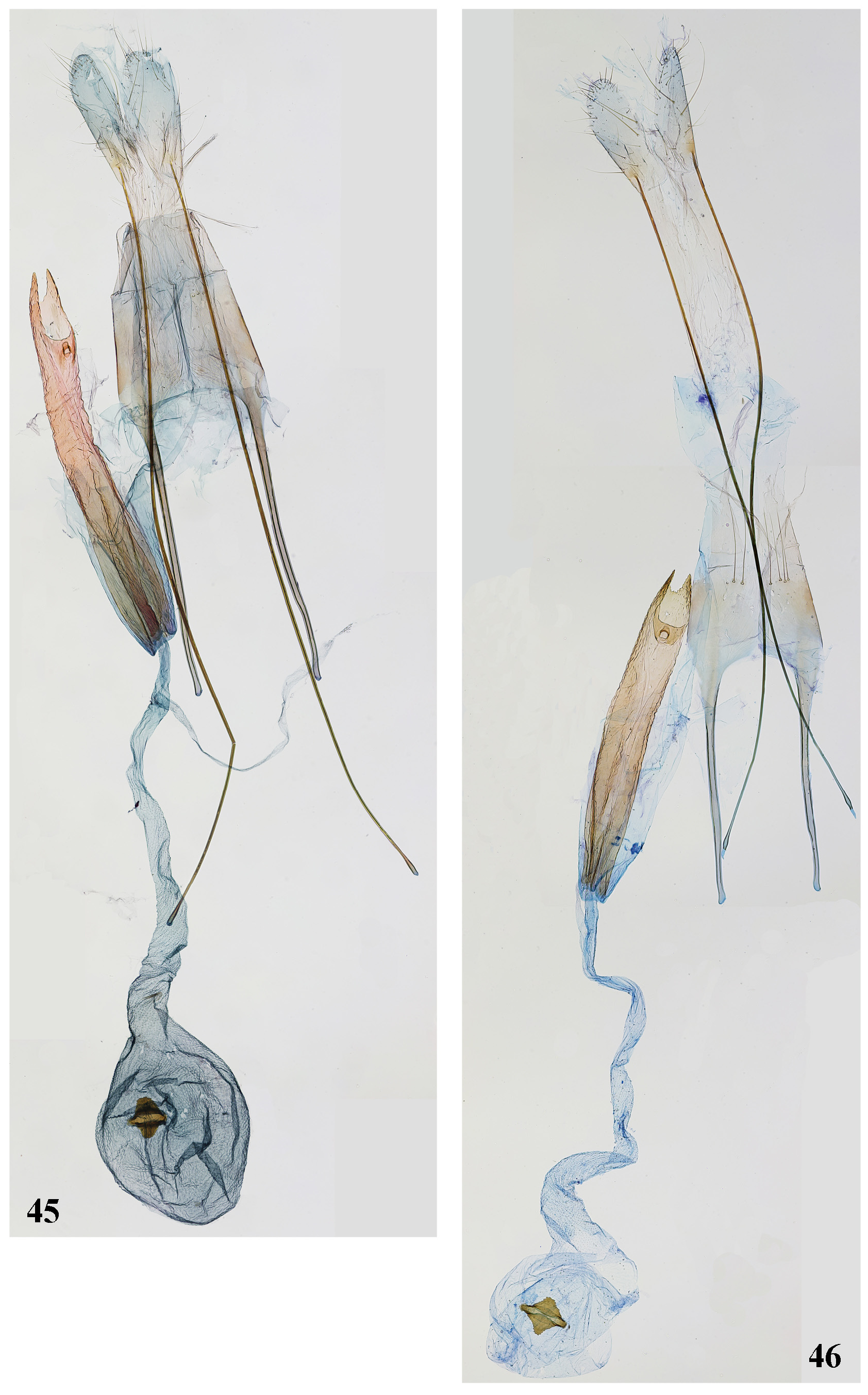
Figs 40–46 View FIGURES 40–44 View FIGURES 45, 46
Type material. Holotype ♀, LepsocAfrica CRG, M1334, Gelechioidea, reared on Ozora obovata var. elliptica , Anacardiaceae , York, Farm, Hoedspruit, South Africa, Limpopo, 24˚15’ 53.4”S, 30˚53’28.5”E, 22.vi.2016 eclosed (A. & I. Sharp) (gen. slide 183/20, O. Bidzilya) ( TMSA). Paratypes: 1♀, LepsocAfrica CRG, M1430, Gelechioidea, reared on Ozora obovata var. elliptica , Anacardiaceae , York, Farm, Hoedspruit, South Africa, Limpopo, 24˚15’53.4”S, 30˚53’28.5”E, 4.xi.2016 eclosed (A. & I. Sharp) (gen. slide 149/19, O. Bidzilya); 1♂, 1♀, LepsocAfrica CRG, M1334c, Gelechioidea, reared on Ozora sphaerocarpa , Anacardiaceae , York Farm, Hoedspruit, South Africa, Limpopo, 24˚15’53.4”S, 30˚53’28.5”E, 21.vii.2016 eclosed (A. & I. Sharp) (gen. slide 19/ 19♀, 22/ 19♂, O. Bidzilya) (all HSS).
Diagnosis. This new species is characterized externally by black-greyish forewing with distinct ochreousbrown markings and light-brown irroration. Other species of Teleiopsis Sattler, 1960 with somewhat similar wing pattern ( T. lunariella (Walsingham, 1908) , T. latisacculus Pitkin, 1988 , T. sophistica (Meyrick, 1935)) have light grey to dark brow rather than ochreous-brown markings along costal margin. The long male abdominal terga VIII (extended beyond coremata), basally broad sacculus extended beyond valva, phallus longer than combined tegumen-uncus, short (shorter than phallus) straight part of ductus ejaculatorius, female with long antrum and ductus bursae entering posteriorly on the corpus bursae justify placement of new species in the diffinis species-group ( Pitkin 1988: 173). The rounded hump before middle on outer margin of sacculus in the male genitalia, and the antrum with large sub-rectangular posterodorsal emargination and with subapical fold bearing a distinct rounded opening in the female genitalia separate T. sharporum sp. nov. from related species.
Description. Adult ( Figs 40, 41 View FIGURES 40–44 ). Wingspan 14.0–16.0 mm. Head covered with brown, apically black scales with light grey tips, frons lighter with brown grey-tipped scales, labial palpus up-curved, palpomere 2 black rarely mixed with white- or brown- to pink-tipped scales on outer and upper surface, and densely mixed with white to light brown on inner surface, lower surface with raised scales and moderately deep medial groove, females with white-tipped scales gathered in diffuse basal and medial rings, palpomere 3 from 2/3 to 1/2 as long and as wide of palpomere 2, acute, black with white or light brown basal and medial ring, apex white; scape black with light grey apex, flagellomeres black brown-ringed, in male pubescent ventrally. Thorax and tegulae covered with brown, apically black scales with light grey tips, forewing covered with grey black-tipped scales diffuse ochreous-brown costal spot just after middle and at 3/4-4/5 length, both costal and dorsal margin, termen and subcostal veins suffused with ochreous to brown and yellowish-brown, paired tuft of raised scales at 1/4, 1/2 and 2/3 length, cilia black grey-tipped; hindwing and cilia light grey. Abdomen grey with tergum IV yellow in male; legs in male black, mixed and ringed with white, hindtibia dorsally with grey dense hairs; legs in female black, distinctly mixed and ringed with white and light brown to pinkish brown, hindtibia dorsally with dense grey or light brow hairs; tergum VIII in male ( Fig. 43 View FIGURES 40–44 ) subtriangular, twice as long as broad at base, basal 1/3 sub-trapezoidal, terminated laterally into short, hump-shaped weakly elongated lobes, distal 2/3 tongue-shaped, gradually narrowed towards weakly rounded apex, anteromedial emargination reverse U-shaped, extending to 1/3 length of sternum VIII; basal coremata composed of long dense tuft of hair-like scales, posterior coremata extending to about 2/3 length of tergum and consisting of numerous strong spiniform pointed scales and several apically hooked scales; sternum VIII ( Fig. 43 View FIGURES 40–44 ) about 2.5 times as broad as long, posterior margin weakly rounded, covered with dense hairs; sternum II in female with additional group of setae near anterior margin; sternum VII ( Fig. 44 View FIGURES 40–44 ) with posterior band of distinct scales bases, extending anteromedially to 1/3 width of sternum. Male genitalia ( Fig. 42 View FIGURES 40–44 ). Uncus narrow, elongate, with short apical hook, covered laterally with short strong setae; gnathos subequal in length to, and slightly broader than uncus, apex weakly rounded; basal half of tegumen equilaterally triangular, anteromedial emargination narrow, reverse V-shaped, distal half of tegumen narrow, parallel-sided, with gradual transition to uncus, with paired strongly sclerotized medial folds; valva slender, needle-shaped, weakly curved, extending to half length of saccullus; sacculus weakly narrowed in distal half, covered with short stout triangular thorns before inwardly curved pointed apex, inner margin of sacculus nearly straight, outer margin with distinct rounded hump before middle; vinculum narrow, posterior projection broadened dorsally; saccus absent; phallus slender, nearly of equal width, extending to the tip of sacculus. Female genitalia ( Figs 45, 46 View FIGURES 45, 46 ). Papillae anales ovate, covered with short setae; apophysis posterioris equaling combined length of ductus bursae and antrum, apophysis anterioris 1/3-1/4 length of apophysis posterioris, straight, weakly broadened at base; segment VIII unmodified, evenly sclerotized, with short anteromedial emargination; antrum weakly as long as apophysis anterioris plus segment VIII, apex deeply emarginated, covered with fine thorns, posterodorsal emargination large, sub-rectangular, sub-apical fold with distinct rounded opening; ductus bursae slender, weakly broadened before corpus bursae, twice as long as antrum, ductus seminalis arising from posterior 1/8; corpus bursae rounded, signum a sub-hexagonal plate, medial ridge narrow, constricted in middle, posterior lobe broader than anterior lobe, serrated.
Biology. The larvae feed on Ozora sphaerocarpa and Ozora obovata var. elliptica (Anacardiaceae) in May-June, and in late October. Adults emerged from late June to late July, and in early November ( Staude et al. 2020: 59, as Teleiopsis sp.).
Distribution. South Africa: Limpopo Province.
Etymology. This species is named in honor of Ian and Allison Sharp in recognition of their contribution to the study of host-plant relationships of South African micromoths.
Remarks. The genus Teleiopsis with Recurvaria diffinis Haworth, 1828 as type species, was introduced by Sattler (1960) for accommodation of R. diffinis and six species from the collective genus Gelechia Hübner, 1825 . As a result of the generic revision ( Pitkin 1988), three additional new species were described, and one species ( T. sophistica ) was transferred to the genus form Telphusa Chambers, 1872 . Later a new species was described from Korea (Ponomarenko & Park 2007), three new species were discovered in Alps ( Schmid 2011; Huemer & Mutanen 2012) and two in the Asian part of Russia ( Bidzilya 2012; Bidzilya & Nupponen 2018). Now 13 species of Teleiopsis are known to occur in the Palaearctic region, with the majority of them in the Mediterranean region. The only Nearctic species is restricted to California. No Teleiopsis species has been recorded in the Afrotropics except for T. lunariella (Walsingham, 1908) which is endemic to the Canary Islands. The current infrageneric classification within Teleiopsis is based on morphological characters: three groups of species are recognized within the genus based mainly on genitalia characters and structure of abdominal segments. The host-plant specialization of larvae corresponds the phylogenetic relationships within the genus as well. Members of the terebinthinella -group feed on Anacardiaceae , whereas species of the diffinis - and brevivalva -groups are associated with Polygonaceae and Juglandaceae respectively ( Pitkin 1988). The placement of the T. sharporum sp. nov. in the diffinis -group of species is well supported morphologically (see above under Diagnosis) despite the fact that its larvae feed on Anacardiaceae . The discovery of this new Teleiopsis in South Africa broadened considerably the range of the genus, and supports the suggestion about restriction of members of this genus to regions with Mediterranean climate and vegetation ( Pitkin 1988: 148, 149).

| TMSA |
Transvaal Museum |
| HSS |
Research Centre of "La Orden-Valdesequera" |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Gelechiinae |
|
Tribe |
Litini |
|
Genus |