
Strongylosomides troglobius Golovatch, Bouzan & Gallo, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5155.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:6E9C5E63-48B6-4D29-B98D-3DA2755A5AB8 |
|
DOI |
https://doi.org/10.5281/zenodo.6669352 |
|
persistent identifier |
https://treatment.plazi.org/id/03AA0A59-BB44-F176-FF48-7521FE43FC4E |
|
treatment provided by |
Plazi |
|
scientific name |
Strongylosomides troglobius Golovatch, Bouzan & Gallo |
| status |
sp. nov. |
Strongylosomides troglobius Golovatch, Bouzan & Gallo View in CoL , sp. nov.
Figs 4–7 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7
Material examined. Holotype male ( LES 0027943 About LES ), Brazil, Bahia State, São Desidério Municipality, Gruta da Baixa Fria cave , aphotic zone, S12°22’55.6”, W44°56’12.0”, 02.XI.2011, M.E. Bichuette, J.E. Gallão, C.S. Fernandes & D.R. Pedroso leg. GoogleMaps
Paratypes: 1 male, 2 females ( LES 0027944 About LES ), same place, taken together with holotype .
Name. To emphasize the cave-dwelling habits of this species, masculine adjective in genitive case.
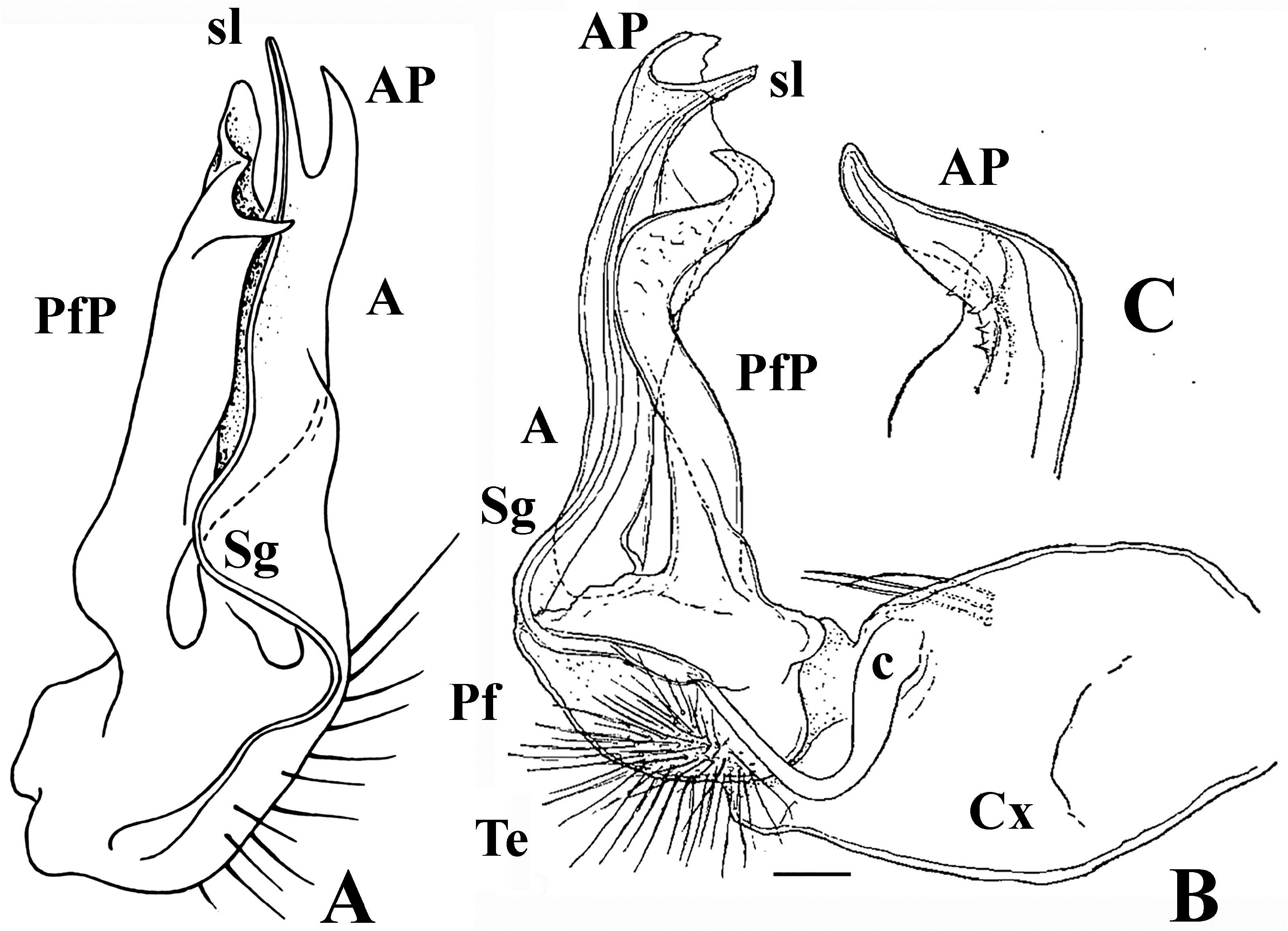
Diagnosis and affinities. This new Strongylosomides species differs clearly from congeners by the light reddish ochre coloration and a rather fragile body with thin tegument (likely troglomorphisms), combined with the absence of modifications on pregonopodal sterna, and of gonocoxal distodorsal apophyses, but above all, by the prefemoral process (PfP) of the gonotelopodite (Te) being fully coalesced in its basal half with the acropodite (A) and leaving the distal half acuminate and directed caudad ( Figs 6 View FIGURE 6 & 7 View FIGURE 7 ). The light ochre body coloration, as well as gonopodal shape and structure of S. troglobius sp. nov. resemble those of S. stercoriarius ( Schubart, 1956) , comb. nov., taken from near a cave in Minas Gerais ( Schubart 1956), so vividly that both these species are undoubtedly very similar. They share the same, rather troglomorphic light ochre coloration, since both have been collected inside or near a cave, respectively. Both share the presence of male tibial chelae as well. More important, however, is that their gonopods are also very similar ( Figs 3A View FIGURE 3 , 6 View FIGURE 6 & 7 View FIGURE 7 ), the gonocoxae (Cx) being devoid of distodorsal apophyses, the acropodites (A) nearly identical, and the prefemoral processes (PfP) long, sinuous and attenuating distad. The only sharp difference concerns the PfP of S. troglobius sp. nov., which is fused by its basal half to A while its distal part is directed caudad, gradually attenuating and strongly sharpened at the end ( Figs 6 View FIGURE 6 & 7 View FIGURE 7 ), vs. the PfP of S. stercoriarius , comb. nov., being completely free from A (as is typical of the genus, Fig. 3B View FIGURE 3 ), ribbon-shaped, clearly sinuous and not so sharp at the end ( Figs 3B & C View FIGURE 3 ).
In other words, we formally propose the following new transfer: Strongylosomides stercoriarius ( Schubart, 1956) , comb. nov. ex Leptodesmus de Saussure, 1859 . Moreover, despite the PfP basal half being conspicuously fused to A in S. troglobius sp. nov., we are inclined to formally assign this species to Strongylosomides , together with and close to S. stercoriarius , comb. nov. As Hoffman (1979) predicted, we can reasonably expect further new species and records of Strongylosomides in the future.
Description. Length of adults ca 15–16 mm (males) or 22 mm (females), width of midbody pro- and metazonae 2.0 and 2.3 mm (males), 3.0 and 3.3 mm or 2.5 and 2.8 mm (females), respectively. Coloration of both live and fixed specimens uniformly light reddish ochre; antennae and legs mostly a little lighter, yellow ochre ( Figs 4 View FIGURE 4 & 5 View FIGURE 5 ).
Body subcylindrical, with 20 rings (18p+1a+T) ( Fig. 4 View FIGURE 4 ). Tegument mainly poorly shining, texture mostly smooth to very delicately striolate. Head densely pilose all over clypeolabral region, vertex almost bare, epicranial suture faint; isthmus between antennae ca 1.2 times broader than diameter of antennal socket ( Fig. 4C View FIGURE 4 ). Mouthparts usual, not modified. Antennae relatively short and slender, poorly clavate due to only a slightly enlarged antennomere 6, in situ projecting past ring 3 dorsally (male) or 2 (female); in length, antennomeres 2 = 6> 3 = 4 = 5> 1> 7 ( Fig. 4 View FIGURE 4 ). Genae regularly rounded laterally ( Figs 4B & C View FIGURE 4 ), incisura lateralis small and round, located beneath antenna; gnathochilarium without peculiarities.
In width, head <collum <ring 2 <3 <4 <5=15 (male, female), thereafter body gradually tapering towards telson ( Fig. 4 View FIGURE 4 ). Dorsum smooth, regularly and strongly convex, at most very faintly rugulose in places, a little more densely and clearly so below paraterga ( Figs 4 View FIGURE 4 & 5 View FIGURE 5 ). Paraterga rather poorly developed, only a little smaller in females, set low, mostly at about midheight of metazonae ( Figs 4 View FIGURE 4 & 5 View FIGURE 5 ), clearly declivous, especially narrowly rimmed on collum, thereafter small, regularly rounded, laterally smooth and anteriorly increasingly incomplete ridges on poreless rings, vs. clearly thickened, also increasingly abbreviated, oblong porostele-like calluses caudally on porebearing rings ( Figs 4 View FIGURE 4 & 5D View FIGURE 5 ). Calluses on paraterga delimited by sulci both dorsally and, in posterior part of body, caudally. Caudal corners of paraterga acutangular and mostly rounded to narrowly rounded, but never drawn past rear tergal margin ( Fig. 4 View FIGURE 4 ). Pore formula normal (5, 7, 9, 10, 12, 13, 15–19); ozopores small, located inside ovoid pits a little before caudal corners of paraterga ( Figs 4 View FIGURE 4 & 5D View FIGURE 5 ). Tergal setae absent, no setation pattern traceable on metaterga. A transverse sulcus or impression on metaterga absent. Stricture between pro- and metazonae distinct, but rather narrow, shallow and smooth. Limbus very thin and entirely smooth. Pleurosternal carinae increasingly low, arcuate, longitudinal ridges or bulges traceable from ring 2 until ring 18 ( Figs 4B, C View FIGURE 4 & 5 View FIGURE 5 ). Epiproct rather long, conical, straight, tip subtruncate; pre-apical papillae small ( Fig. 4 View FIGURE 4 ). Hypoproct roundly subtriangular, sharpened at tip; setigerous papillae at caudal corners small and well separated; sides convex ( Fig. 4C View FIGURE 4 ).
Sterna broad, without notable modifications even before ring 7, nearly glabrous, cross-impressions weak ( Fig. 4C View FIGURE 4 ). Epigynal ridge very low. Legs relatively short and slender ( Figs 4 View FIGURE 4 , 5C & D View FIGURE 5 ), especially so in female, ca 1.3–1.4 (male) or 0.9–1.0 times (female) as long as midbody height; male legs with chelae (distoventral protrusions on tibiae) gradually disappearing towards ring 14; in length, femur >> prefemur = tarsus> tibia> coxa = postfemur; claw very short ( Figs 4 View FIGURE 4 , 5C & D View FIGURE 5 ). Male gonopores located on distinct, distomesal, subtriangular gonapophyses ( Fig. 4C View FIGURE 4 ).
Gonopodal aperture ovoid, simple, only a little broader than following sterna ( Fig. 5C View FIGURE 5 ). Gonopods ( Figs 5A, B, D View FIGURE 5 , 6 View FIGURE 6 & 7 View FIGURE 7 ) stout and robust, devoid of sternal rudiments; gonocoxa (Cx) devoid of a distodorsal apophysis, cannula (c) as usual, a thin curved tube, devoid of setae mesally at its base; prefemoral region (Pf) as usual, short, about half as long as an untwisted acropodite (A); caudal prefemoral process (PfP) long, gradually attenuating distad, fused by its basal half to A, its distal half directed caudad, attenuating and strongly sharpened apically ( Figs 6 View FIGURE 6 & 7 View FIGURE 7 ); A laminar and membranous, suberect; seminal groove (Sg) running entirely on mesal side of telopodite (Te), ending up on a short solenomere (sl) opposite a similarly short end branch, or acropodital process (AP) ( Fig. 6 View FIGURE 6 ).
Vulvae not studied: one female paratype dissected, but no vulvae found (?underdeveloped).
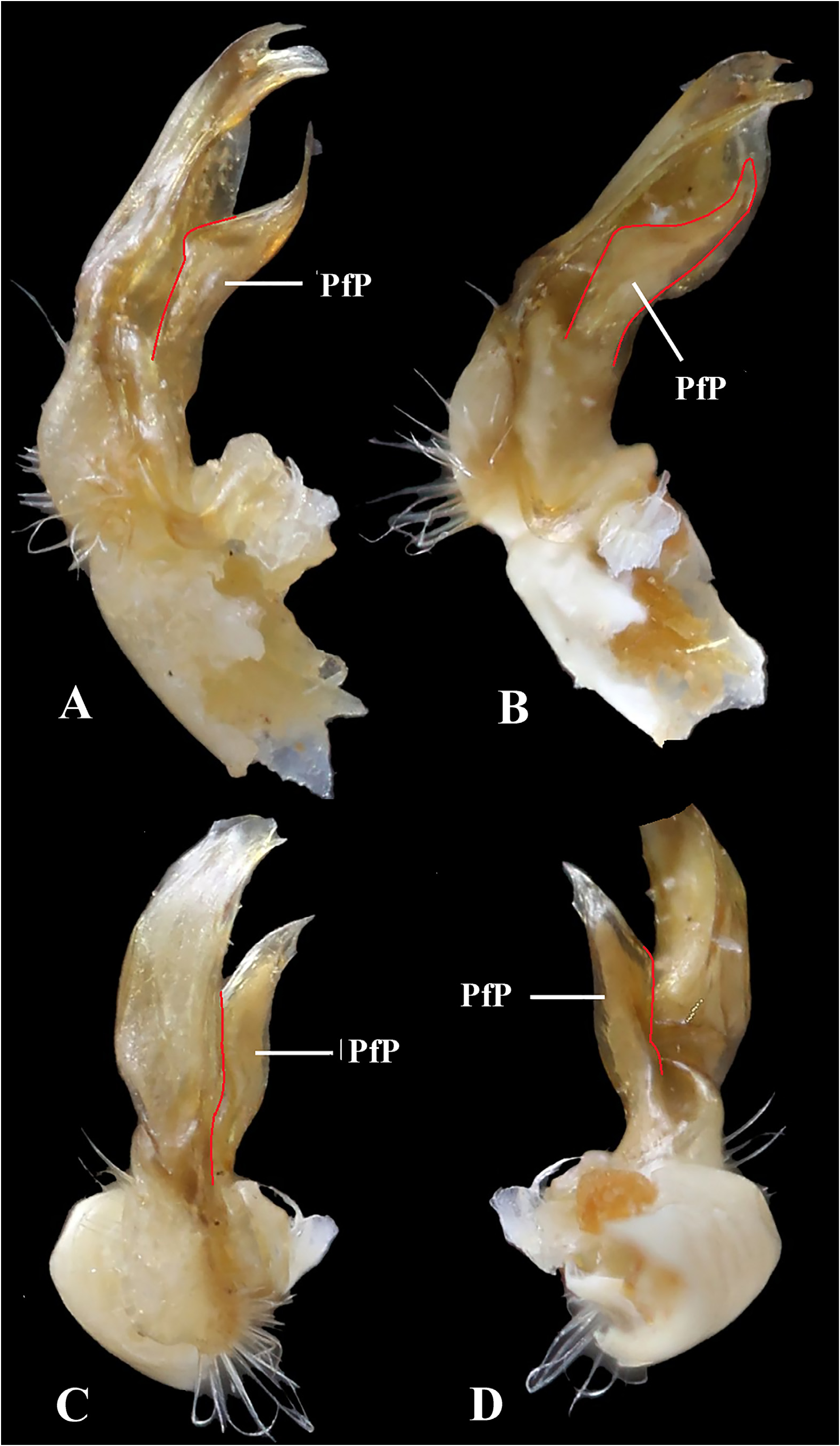
Remarks. Superficially, the gonopodal telopodite of S. troglobius sp. nov. looks uniramous at the base, because the basal half of the prefemoral process (PfP) is tightly appressed to and coalesced with the acropodite (A), the latter branching off at about midway and independent only distally ( Fig. 6 View FIGURE 6 ). However, a closer examination of the PfP shows clear traces ( Fig. 7 View FIGURE 7 , outlined in red) of the fusion of the PfP base to the acropodite base in all meaningful aspects. This betrays the basic bipartite condition typical of the Chelodesminae gonopod, including Strongylosomides as a genus that remains unassigned to a tribe yet.
The absence of a PfP is characteristic of the Prepodesminae ( Hoffman & Reid 1990) and several genera of Chelodesminae , both with a definite tribal position (e.g., Telonychopodini) and still not classified in accepted tribes. Among the latter genera, such are, e.g., Cryptosolenomeris Hoffman, 2009 , monobasic, with one species from Espírito Santo state, Brazil ( Hoffman 2009), or Odontopeltis Pocock, 1894 , whose eight presently known species, all in southeastern Brazil (Espírito Santo, Minas Gerais and Rio de Janeiro) ( Hoffman 1981; Pena-Barbosa et al. 2013), are among the largest in the family (80–107 mm long). However, none of them show a structure resembling a PfP, even such that is fused with discernible traces to the base of the gonopodal acropodite (A).
Because both S. troglobius sp. nov. and S. stercoriarius , comb. nov., differ from the remaining congeners by the rather pale coloration (vs. dark), the unmodified pregonopodal sternites 4, 5 and 6 (vs. with paramedian cones), the gonocoxae devoid of distodorsal apophyses (vs. present), the basal course of the seminal groove running closer to the ventral margin of the acropodite (vs. abruptly reflexed dorsad across the prefemoral region and proceeding to the solenomere along the dorsomedian edge of the acropodite), the prefemoral process attenuating, directed caudomesad and sharp at the end (vs. broad and suberect), and the absence of a midway cingulum on the acropodite (vs. often present) (cf. Fig. 3A View FIGURE 3 and Fig. 3B View FIGURE 3 ), another separate species-group of Strongylosomides , the stercoriarius - group, seems warranted to encompass both S. troglobius sp. nov. and S. stercoriarius , comb. nov. The latter species thereby clearly bridges S. troglobius sp. nov. to the remaining congeners.
The few supposedly troglomorphic traits of S. troglobius sp. nov. alone, the more so as the mouthparts being unmodified, as is sometimes the case in troglobionts, and the antennae and legs relatively short, are certainly not enough for conferring the species the status of a troglobiont. Only direct biological observations will allow for this to be evaluated unequivocally. Even though epigean sampling was conducted on the same day near the cave entrance, with no additional material of S. troglobius sp. nov. revealed, this species was only found inside the cave in the deepest zones. Isolation in the hypogean environment, combined with certain presumably troglomorphic traits such as reduced body pigmentation and a quite fragile body with thin teguments, seems to be evidence of a neotroglobiont status of this species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |