
Satellitenemertes satellitensis, Iwata, 2006
|
publication ID |
https://doi.org/ 10.1080/00222930600833800 |
|
DOI |
https://doi.org/10.5281/zenodo.4672286 |
|
persistent identifier |
https://treatment.plazi.org/id/573F4468-FFC9-FFC4-FEA7-F475220FFA74 |
|
treatment provided by |
Carolina |
|
scientific name |
Satellitenemertes satellitensis |
| status |
|
Satellitenemertes satellitensis gen., sp. nov.
Type specimen
Holotype: USNM 1072179 About USNM ; 15 slides of stained serial section, including transverse sections of the anterior portion of the body (8) and proboscis (5) and horizontal sections of the proboscis (2). Two short pieces were not sectioned.
Type locality
Satellite Channel , Vancouver Island, British Columbia, Canada (Lat 48 ° 41.9 9 N, Long. 123 ° 28 9 W), depth 15–22 fathoms, September 9, 1964 .
Description
External features
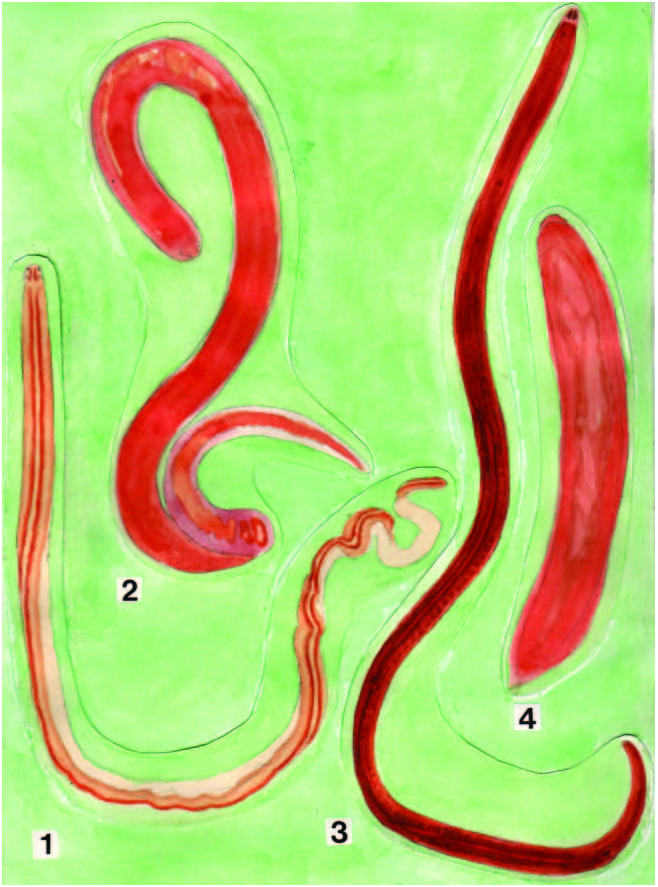
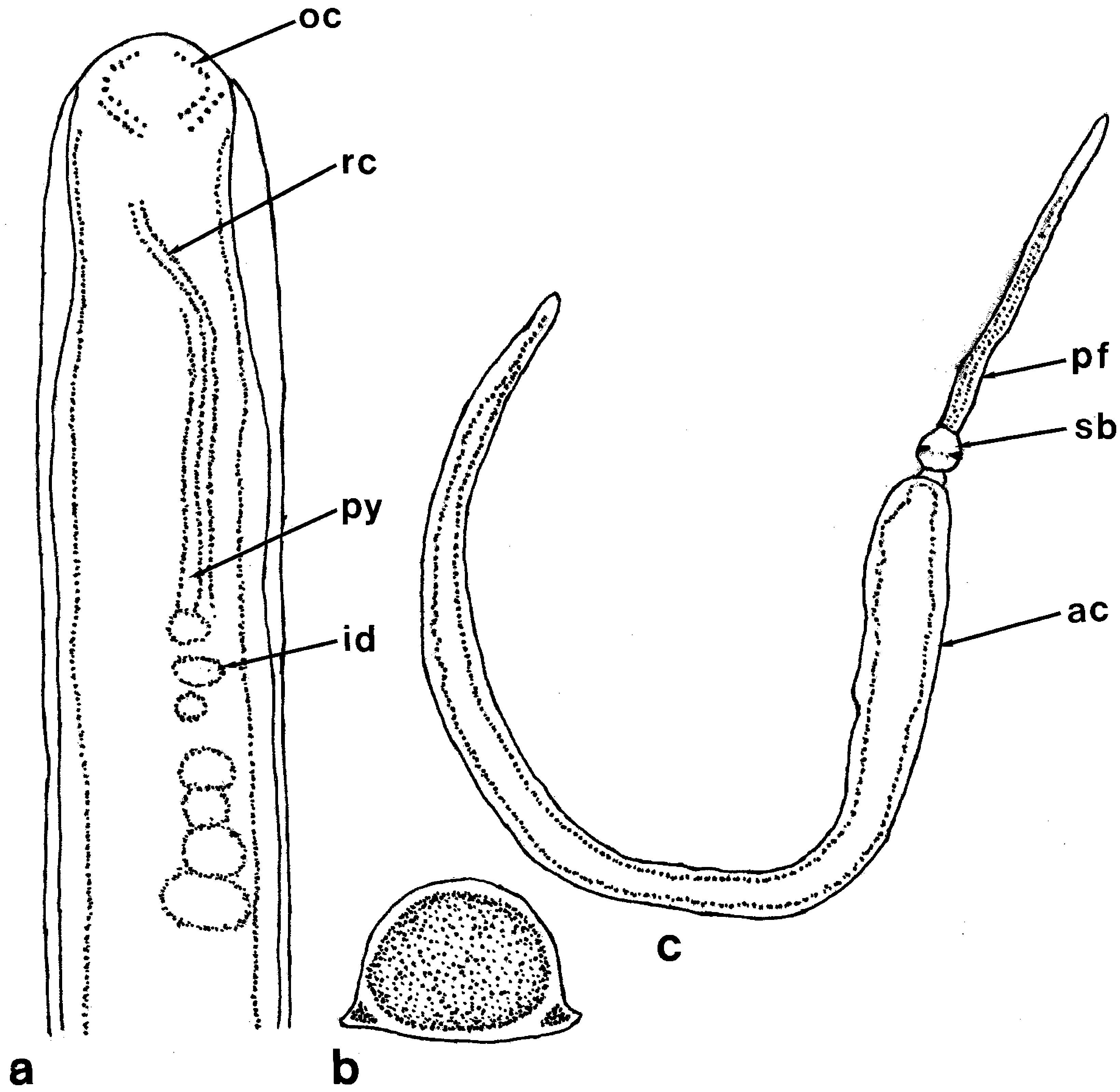
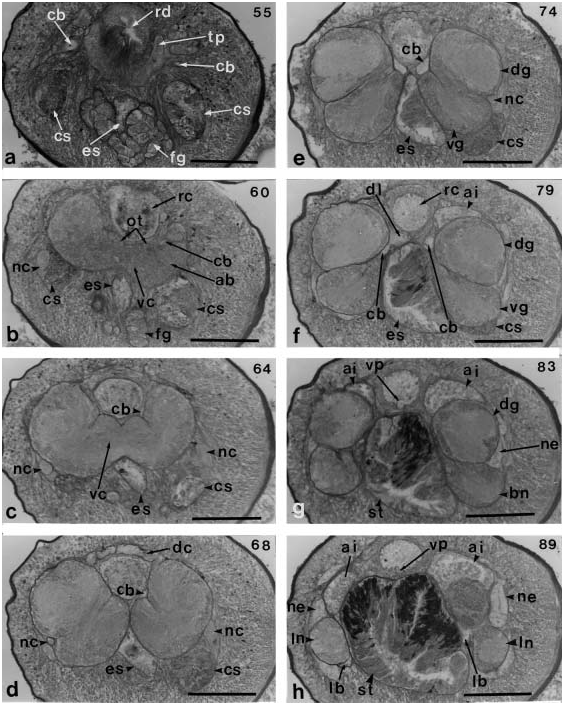
The worm was small and nearly cylindrical, 2 cm long and 0.8 mm wide, and moved slowly ( Figure 1.2 View Figure 1 ). The anterior end of the head was rounded when viewed from above ( Figure 7a View Figure 7 ). Neither cephalic grooves nor the proboscis pore were observed. The general color of the body was orange, but the lengthwise ventrolateral ridges of the body wall were more brightly coloured ( Figure 7b View Figure 7 ). There were four groups of ocelli. Each of the two more anterior-most groups consisted of about ten ocelli arranged in a nearly semicircular pattern; behind these were two groups consisting of five or six ocelli. The proboscis sheath was visible through the body wall as a slender duct in the anterior portion of the body ( Figure 7a View Figure 7 ). Two days after the worm was captured, the proboscis was protruded. It was long, stout and reddish orange in color; three portions (anterior and posterior chambers, proboscis diaphragm and stylet bulb) were easily recognized ( Figure 7c View Figure 7 ).
Body aeall, musculature and parenchyma
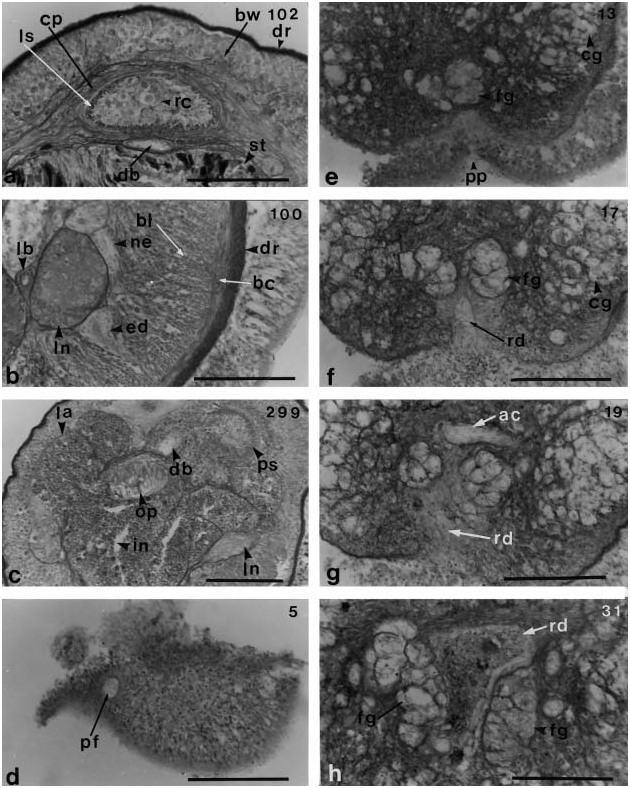
The epidermis of the dorsal side was damaged during fixation ( Figure 8a View Figure 8 ). It contains unicellular elliptical glands sparsely arranged in a row, and there are no cyanophilic glands between them ( Figure 8b View Figure 8 ). The dermis is thin, averaging 7–15 mm in thickness ( Figure 8a, b View Figure 8 ). The body wall musculature is weakly developed and has a thin and sparse layer of lattice-type diagonal muscles between the outer circular and inner longitudinal muscle layers ( Figure 8b View Figure 8 ). The outer circular muscle layer, 10 mm in thickness, is thin, while the inner longitudinal muscle layer is 90 mm thick. The longitudinal muscle layer is not divided anteriorly by connective tissue and cephalic retractor muscles are absent. Dorsoventral muscles and parenchyma are also absent ( Figure 8c View Figure 8 ).
Rhynchodeum, rhynchocoel and proboscis
The pre-cerebral region lacks a short snout or a mid-ventral epidermal furrow. The proboscis pore is a short median ventral groove (30 mm long), 50 mm behind the opening of the frontal organ ( Figure 8d, e View Figure 8 ). The rhynchodeum rises upward behind the transverse anastomoses of the cephalic blood vessels, is bordered by a pair of large frontal glands, and then extends to the dorsal side of these ( Figures 8 View Figure 8 f–h and 9a). It is covered by a thick layer of circular muscles (20–40 mm) and opens ventrally into the oesophagus ( Figure 9b, c View Figure 9 ). The rhynchodeum is provided with a thick epithelium folded internally ( Figure 9d View Figure 9 ). The precerebral septum of the proboscis apparatus is of the closed type, and composed of radial bundles of muscles from the longitudinal muscle layer of the body wall ( Figure 9e View Figure 9 ).
The rhynchocoel has two distinct muscle layers and reaches posteriorly more than half the body length ( Figure 8a View Figure 8 ). The anterior region of the proboscis is provided with outer circular and inner longitudinal muscle layers. The proboscis insertion could not be observed because the proboscis had been protruded.
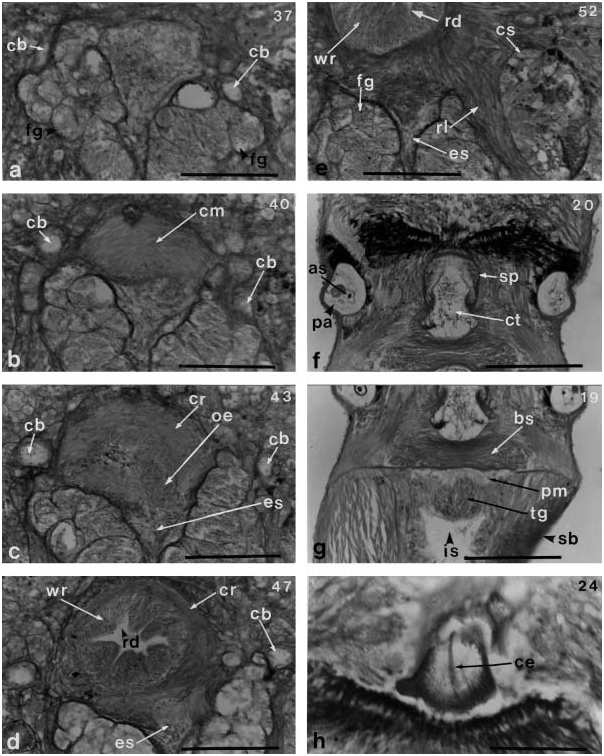
The proboscis diaphragm has a central stylet apparatus consisting of a pear-shaped basis 90 mm long and 60 mm wide; the anterior half of the basis is enclosed by a sheath ( Figure 9f, g View Figure 9 ). The central stylet is 39 mm long ( Figure 9h View Figure 9 ). The basis has no posteriorly inserted accessory stylet. It rests on a large bolster, 150 mm wide and 30 mm thick, consisting of interwoven circular and longitudinal muscle ( Figure 9f, g View Figure 9 ). The stylet bulb, 110 mm wide and 30 mm high, consists of plate-like connective tissue and has a thick layer of glandular cells in anterior portion of its wall ( Figure 9g View Figure 9 ). The remainder of the wall has a layer of connective tissue and a narrow canal into the posterior chamber of the proboscis. There are two accessory stylet pouches, each containing two stylets ( Figure 9f View Figure 9 ). The proboscis has 11 nerves.
Alimentary canal
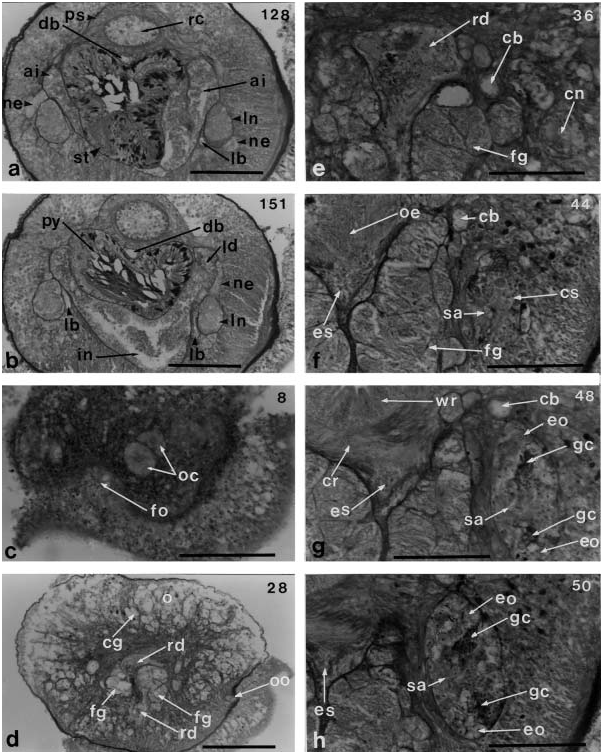
The foregut opens into the rhynchodeum near the anterior end of the brain ( Figure 9c View Figure 9 ). The anterior part of the oesophagus is bordered on both sides by large masses of frontal glands ( Figures 9 View Figure 9 a–e and 10a), and posteriorly it is surrounded by longitudinal muscles ( Figure 10b View Figure 10 ). It has a length of 140 mm, measured from the opening of the oesophagus to the anterior end of the brain, and then extends for 50 mm under the ventral commissure of the brain ( Figure 10c View Figure 10 ). The transition of the oesophagus into the stomach, measuring 90 mm long, is found between the brain lobes ( Figure 10 View Figure 10 d–f).
The stomach (330 mm long) has four deep folds, is longer than the brain (250 mm) and lacks a diverticulum ( Figure 10g, h View Figure 10 ). The pylorus is four times as long as the stomach, and its posterior end opens into the intestine ( Figures 8c View Figure 8 and 11a, b View Figure 11 ). The intestinal caecum has a pair of anterior diverticula, 460 mm long, and two pairs of lateral diverticula, which begin at the level of the posterior portion of the brain ( Figures 10 View Figure 10 f–h and 11a, b). The intestinal canal has deep lateral diverticula ( Figure 8c View Figure 8 ).
Blood υascular system
The blood vascular system has three longitudinal vessels. The two cephalic blood vessels anastomose above the rhynchodeum near the tip of the head to form a simple vascular loop ( Figure 8g View Figure 8 ). Farther posteriorly, the cephalic vessels do not become large lacunae, but enter the brain ring without giving off cerebral vessels and run alongside the rhynchocoel ( Figure 10e View Figure 10 ). The dorsal vessel originates from the left cephalic vessel and enters the rhynchocoel wall where it forms a median vascular plug ( Figure 10f, g View Figure 10 ). It extends about 120 mm mid-ventrally in the sheath to anterior stomach region. After passing ventrally out of the rhynchocoel, the dorsal vessel continues posteriorly under the proboscis sheath between the rhynchocoel and the alimentary canal. The three post-cerebral vessels are not transversely linked by pseudometameric connectives. In the posterior portion of the brain, the lateral vessels enter the nephridial region but have no loops in contact with the nephridial tubes ( Figure 10h View Figure 10 ).
Nerυous system
The brain and lateral nerves are covered by a thin layer of fibrous connective tissue. ( Figures 8b View Figure 8 and 10 View Figure 10 c–h). The cerebral ganglia are large, but contain no neurochord cells and lack an inner neurilemma. The dorsal ganglia do not have bifurcated fibre cores ( Figure 10h View Figure 10 ). The right and left ganglia are connected by a short dorsal commissure 20 mm thick and a short ventral commissure 60 mm thick, the latter a little more anterior than the former ( Figure 10 View Figure 10 b– d). The ventral ganglia are separated from the dorsal ganglia posteriorly ( Figure 10g, h View Figure 10 ). The lateral nerve cords are without accessory nerves, myofibrillae, or peripheral nerves ( Figures 8b View Figure 8 and 10h View Figure 10 ). The root of the proboscis nerves originating from the right and left dorsal sides of the ventral commissure, becomes a U-shaped trunk, from which the proboscis nerves extend toward the rhynchocoel ( Figures 10a, b View Figure 10 and 12 View Figure 12 e–g). There is no mid-dorsal nerve. A pair of the foregut nerves originates from the ventral ganglia immediately behind the ventral commissure and a transverse connective.
Special sensory organs and frontal organ
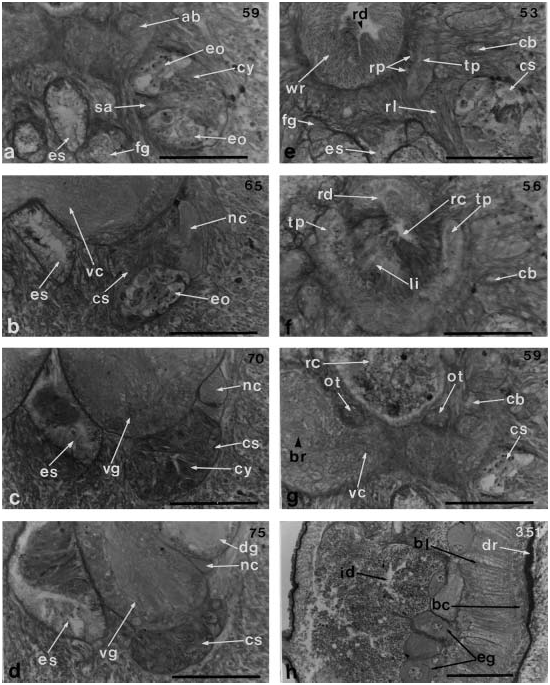
The ocelli in cross-section are found as spherical masses of large ocular cells, the inner surfaces of which form small pores ( Figure 11c View Figure 11 ). Four ocelli are clearly arranged in the right side of the tip of the head and the other ocelli are not detectable among the cephalic glands. The frontal organ opens on the ventral side of the anterior end of the head ( Figure 8d View Figure 8 ). The frontal glands extend posteriorly as a pair of large ventrolateral masses that reach the anterior end of the cerebral ganglia ( Figure 10 View Figure 10 a–c). The largest portion of the mass is 200 mm high and 80 mm wide at the mouth region, which is 350 mm from the tip of the head. The cephalic glands are well-developed on the lateral sides of the head and extend posteriorly. The bases of these cells are situated in the following body regions: close to the opening of the oesophagus, 150mm from the proboscis pore or 130 mm anterior to the brain ( Figure 9c View Figure 9 ). Sub-muscular glands are absent. There are no cephalic grooves ( Figure 11d View Figure 11 ).
The cerebral sensory organs are large 310 mm long and reach below the brain lobes, but do not extend behind them as they do in the members of Nipponnemertes ( Figure 10 View Figure 10 e–g). The cerebral organ canal (110mm long) is found on the lateral side of the head, near the anterior tip, and the lateral sac is not divided at its posterior end ( Figure 11e, f View Figure 11 ). The sensory canal of the cerebral sensory organ, 180 mm long by 90 mm wide at its largest portion, is located medially. The cerebral organ is provided with large eosinophilic glands and ganglionic masses on both the dorsal and ventral sides ( Figure 11g, h View Figure 11 ). Behind the sensory canal, a mass of cyanophilic glands run underneath the brain until its posterior portion ( Figures 10 View Figure 10 b–f and 12a–d). A thick nerve (100 mm long) derived from the dorsal ganglion enters the posterior portion of the cerebral sensory organ ( Figures 10 View Figure 10 c–e and 12b–d).
Excretory and reproductiυe systems
The excretory tubules are weakly developed and extend from the level of the posterior portion of the brain or of the posterior end of the cerebral sensory organ to the end of the pylorus. The two efferent ducts, originating in the anterior portion of the nephridial region, are short; they are lateral to the nerve cords and open on the lateral side of the body ( Figure 8b View Figure 8 ). Sexes are separate. The eggs are arranged individually in a row between the body wall musculature and the intestine ( Figure 12h View Figure 12 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |