
Rhynchelmis (Sutroa) diespluviae Fend
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3760.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:587163F9-326C-4CBC-94A5-4605C8ADB000 |
|
DOI |
https://doi.org/10.5281/zenodo.6139330 |
|
persistent identifier |
https://treatment.plazi.org/id/0397878B-FF85-FFB1-FF4D-5CE2FAC4FF6A |
|
treatment provided by |
Plazi |
|
scientific name |
Rhynchelmis (Sutroa) diespluviae Fend |
| status |
sp. nov. |
Rhynchelmis (Sutroa) diespluviae Fend View in CoL n. sp.
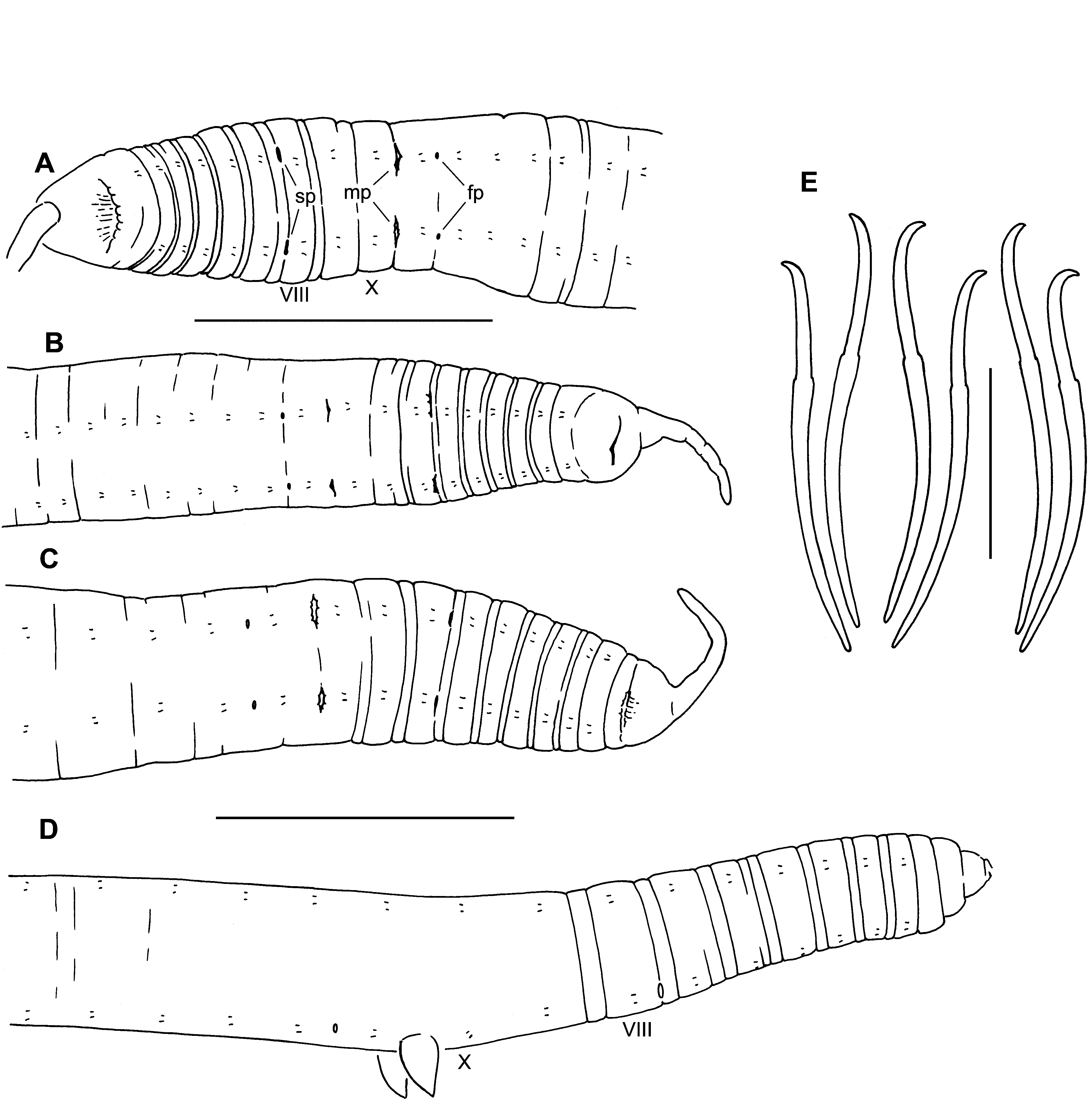
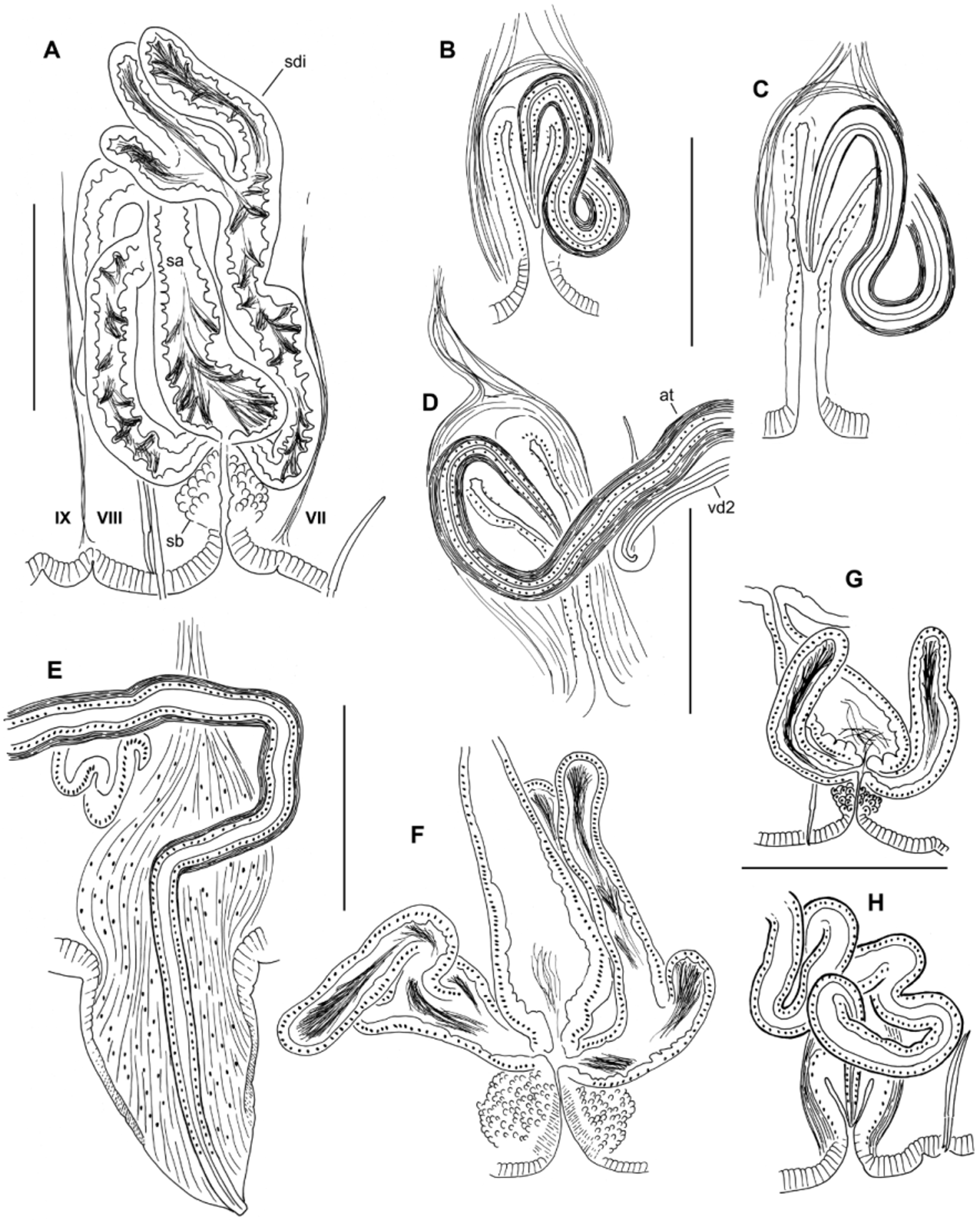
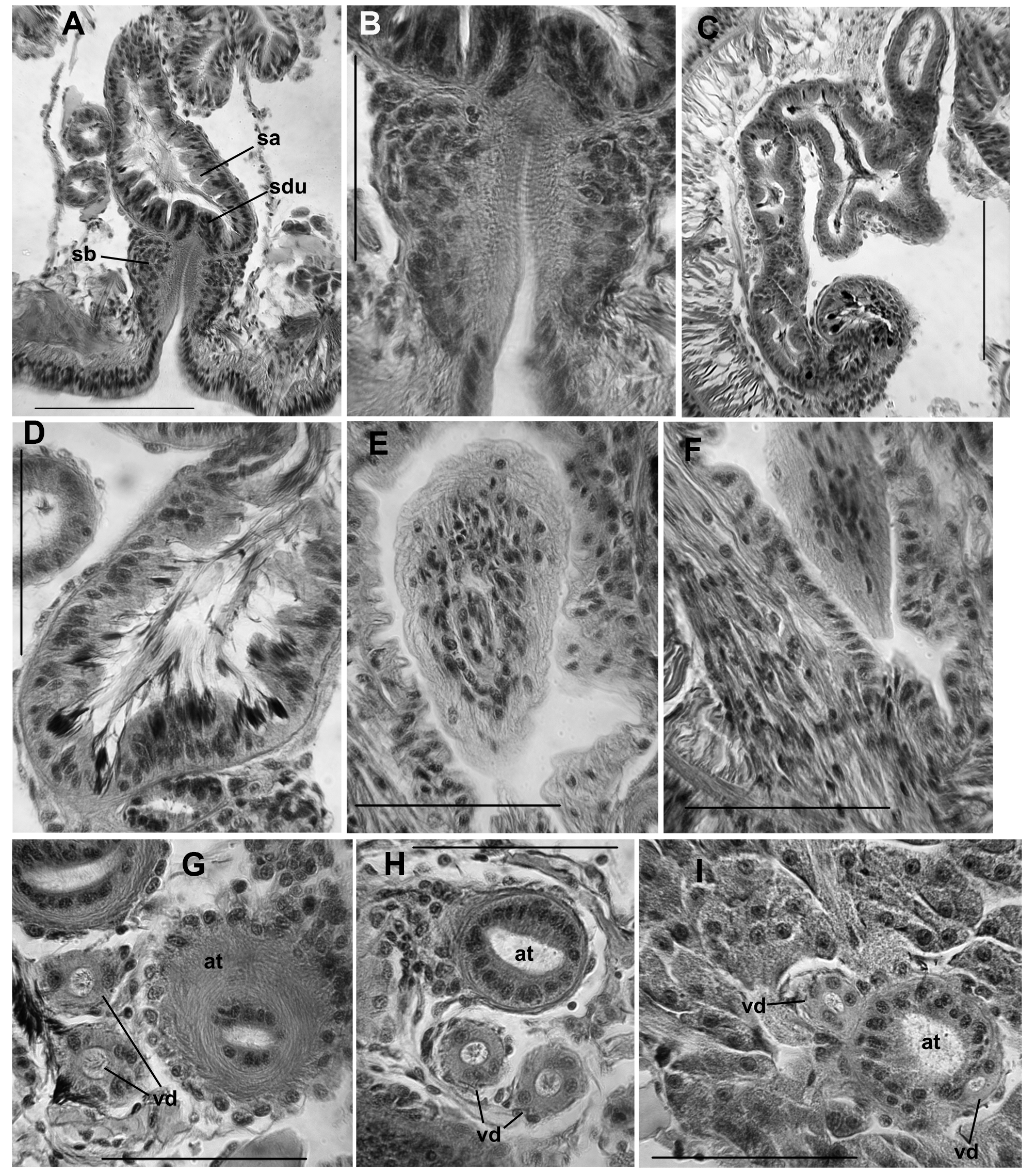
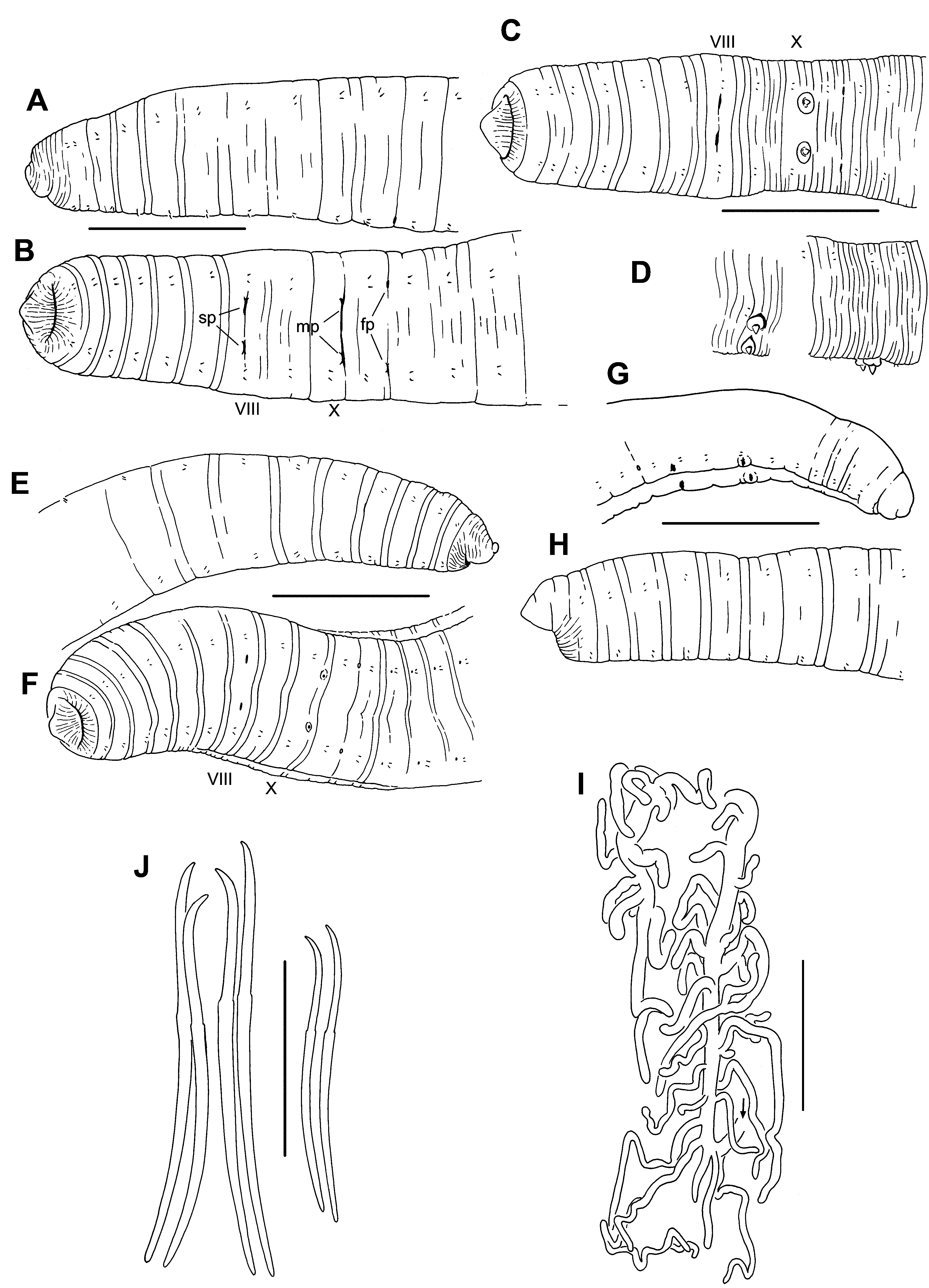
( Figs 6–9 View FIGURE 6. R View FIGURE 7. R View FIGURE 8. R View FIGURE 9. R )
Holotype. USNM 1230311, a dissected, slide-mounted specimen.
Type locality. IDAHO, Idaho Co: Rainy Day Creek at mouth, N45.8030 °, W115.6940 °, 23 Apr 2001.
Paratypes. USNM 1230312-1230314, the type locality, 23 Apr 2001, 2 dissected, 2 sectioned (1 transverse, 1 sagittal). CASIZ 192766, the type locality, 23 Apr 2001, 3 dissected.
Other material. IDAHO, Idaho Co: Rainy Day Creek at mouth, 23 Apr 2001, 12 dissected, 2 in alcohol; 1 Jul 2002, 4 dissected, 3 in alcohol. O’Hara Creek at Selway River, 23 Apr 2001, 1 dissected. Clearwater Co.: Cold Spring Creek at mouth, 2 Jul 2002, 1 dissected. Fern Creek at Isabella Creek, 28 Jul 2003, 2 dissected. North Fork Clearwater River at Isabella Creek, 28 Jul 2003, 2 dissected (partially mature). North Fork Clearwater River at Cedars, 3 Jul 2002, 3 dissected. Orogrande Creek tributary, 2 Jul 2002, 2 dissected. WASHINGTON, Jefferson Co.: Hurst Creek at Clearwater River, N47.5733 °, W124.2917 °, 14 Apr 2010, 1 dissected. All Idaho specimens collected by D.L. Gustafson; the Washington specimen collected by S.V. Fend.
Etymology. The specific epithet refers to the type locality, Rainy Day Creek; from Latin “dies pluviae”.
Description. Idaho material. Length of 6 intact, preserved specimens 21–36 mm; 92–130 segments. Width 0.79–1.35 mm in X (n = 14), maximum width to 1.4 mm. Prostomium with a filiform proboscis, length 0.6–1.1 mm ( Fig. 6 View FIGURE 6. R A–C); body nearly round in transverse section, tapered posteriorly. External segmentation moderately developed; segments III–IX usually with a secondary annulation at about the anterior 1/4; external segmentation obscure in clitellum and weak posteriorly. Clitellum usually from mid-IX to mid-XIV. Chaetae narrowly paired, with bundles approximately equidistant. Chaetae simple-pointed, sigmoid, with a moderately developed distal hook; nodulus distal to midpoint, 28–38 % of chaeta length from tip ( Fig. 6 View FIGURE 6. R E). Chaeta length 151–247 (203) µm in middle segments, similar or slightly smaller in posterior segments (166–206 Μm); dorsal pairs about as long as ventrals. Paired spermathecal openings distinctly anterior to ventral chaetae, on the furrow marking the anterior secondary annulus; transverse position in line with or slightly lateral to chaetae ( Fig. 6 View FIGURE 6. R A–C). Male pores posterior to ventral chaetae in X; transverse position on chaetal line. Female pores at 11/12, in line with the ventral chaetae.
Epidermis 18–36 µm thick in anterior segments, maximum thickness 48–72 µm thick in clitellum. Body wall longitudinal muscle layer 40–70 µm thick in preclitellar segments; circular muscle layer 6–14 µm. Pharynx thickened, with columnar cells in II (dorsally) and III (ventrally) through IV, without a distinct dorsal pad. Pharyngeal glands in IV–VI(VII).
Ventral blood vessel divides at 6/7 or in VII. A pair of convoluted lateral blood vessels joins the dorsal and ventral blood vessels in posterior part of anterior segments; beginning about IX, there may also be an anterior pair of lateral vessels. In X and XI the posterior (commissural) vessels loop into the sperm and egg sacs, respectively. Posterior to about XII, lateral vessels may be blind, or may join the perivisceral sinus on the ventrolateral aspect of the gut; branching is variable, and sometimes not apparent. In a few segments, beginning in VIII or IX, 2–3 small blood glands at the perivisceral sinus are joined by short branches from the ventral blood vessel. Lateral blood vessels indistinct in most specimens, and may be weakly developed or absent in posterior segments; when visible, only one pair is usually well developed.
Nephridia usually from XIII, paired, single, or absent in posterior segments. A small anteseptal funnel is followed by an ovate, granular, postseptal mass (about 150–250 Μm long), which narrows to a tubule; the tubule forms a closely paired loop, which closely follows the anterior lateral blood vessel dorsally, to approach the dorsal blood vessel; this part of the tubule densely covered by granular cells. The tubule loop may then extend ventromedially within the segment. The ectal end of the tubule forms a thin extension to an inconspicuous nephropore, anterior to the ventral chaetal bundle.
Testes paired in X only. Testes and ovaries relatively small, usually ending before mid-segment. Female funnels with posterior lip about 180 Μm high. Sperm sacs extend posteriorly from X to XXV–XXXVI in mature worms; egg sacs extend posteriorly through 1–5 additional segments; no anterior sperm sacs. Paired septal cell masses, probably vestigial sacs, extend posteriorly from 8/9 and 9/10, into IX and X, usually as far as mid-segment.
Spermathecae paired in VIII. External pore formed by invagination of the epidermis, which is lined with a fibrous layer and surrounded by a bulb-shaped mass of densely packed cells, 95–180 Μm high and 100–157 Μm wide; bulb cells are not obviously glandular ( Fig. 9 View FIGURE 9. R B); no accessory glands at pore. Spermathecal ampulla pyriform, 290–500 Μm long, maximum width at base 120–170 Μm, narrowing to join the ventrolateral side of the gut ( Figs 7 View FIGURE 7. R B, 8A, 9A). One pair of diverticula inserts at the junction between bulb and ampulla, on anterior and posterior side ( Fig. 8 View FIGURE 8. R A); ectal duct very short and indistinct ( Fig. 9 View FIGURE 9. R A, B). Spermathecal diverticula roughly tubular, but usually with 1–3 branches ( Figs 8 View FIGURE 8. R A, 9C), usually longer than the ampulla, 490–960 µm long by about 60–90 Μm in diameter. Muscle layer surrounding ampulla and diverticula thin and indistinct, 1–2 Μm thick. Epithelium of diverticula and ampulla may be irregularly incised or folded, especially near base; sperm concentrated in base of ampulla and throughout the diverticula, with heads lined up along epithelium in the basal half, often concentrated in the epithelial folds ( Fig. 9 View FIGURE 9. R D).
Elongate-conical penes are retracted into deep sacs in all Idaho specimens examined; penes slightly curved mediad, length 110–215 Μm, width at base 55–175 Μm ( Figs 7 View FIGURE 7. R C, 8B–D, 9E–F). Penial sacs are lined with a thick, folded epithelium, surrounded by a diffuse mass of tissue which appears to be mostly muscle fibers; the entire structure about 300–500 Μm high, general form resembling the penial bulbs of other R. ( Sutroa ) species. Ectally, the muscle layer is continuous with the circular muscle layer of the body wall; entally the muscle fibers coalesce to form retractor muscles, joined to the dorsolateral body wall ( Fig. 7 View FIGURE 7. R C). Ectal end of atrium narrows, crosses medial face of penial bulb, then enters bulb from anterodorsal side ( Figs 7 View FIGURE 7. R C, 8B–D); atrial duct continues to narrow, but remains visible within the penis ( Fig. 9 View FIGURE 9. R E); ectal pore at the tip of the penis ( Fig. 9 View FIGURE 9. R F). Atria tubular, 1850–4390 Μm long, extending straight back or sometimes highly contorted, extending posteriorly to as far as XX (median XVIII) when straight. Atria about equally divided into an ectal, muscular part, and an ental, prostate-bearing part ( Fig. 7 View FIGURE 7. R A). Ectal part 50–84 Μm in diameter, with muscle coat 8–14 Μm thick, and cuboidal epithelium about 7–14 thick near the male pore ( Fig. 9 View FIGURE 9. R G); muscle becoming thinner entally, to 2–4 Μm ( Fig. 9 View FIGURE 9. R H). Muscle layer of mostly transverse-circular fibers, with a very thin outer layer of longitudinal muscles. Ental part of atrium 48–72 Μm in diameter, muscle layer 2–4 Μm thick, epithelium 14–20 Μm thick, covered with petiolate, multicellular prostate glands ( Fig. 9 View FIGURE 9. R I). Prostate glands granular, about 100–170 Μm long. Anterior male funnels cup-shaped, to about 120 Μm high, without sperm; anterior (nonfunctional) vasa deferentia 20–30 Μm thick. Posterior male funnels with sperm; highly convoluted, about 250 Μm high, on 10/11; posterior vasa deferentia 24–37 Μm in diameter. Vasa deferentia ciliated, not obviously glandular. Both vasa deferentia are free within the sperm sac adjacent to the ectal part of the atrium ( Figs 7 View FIGURE 7. R A, 9G–H), and join the atria within the prostate-covered, ental portion ( Fig. 9 View FIGURE 9. R I). One vas joins near the beginning of the prostate layer, travels a short distance under the muscle coat, then enters atrial lumen; the other travels under the atrial muscle coat for most of its length, entering near the apex (ental end).
Washington specimen. Width in X and maximum width 1.1 mm, tail and proboscis broken. Clitellum IX– XIV. Chaeta length 192–274 µm in middle segments, nodulus 30–41 % of chaeta length from tip. Spermathecal pores and blood vessels as described for typical (Idaho) specimens. Sperm sacs to XVIII, eggs in XIX; testes only in X. Spermathecae as described for typical specimens; ectal bulb 190 Μm high and about 200 Μm wide. Spermathecal ampulla narrowly pyriform, 410–440 Μm long, maximum width at base 155–170 Μm; diverticula 430–560 µm long, with 2–3 branches ( Fig. 8 View FIGURE 8. R F). Both penes everted, narrowly conical, directed slightly anteriad; length 385–395 Μm, width near base 250 Μm; male pore terminal. ( Figs 6 View FIGURE 6. R D, 8E). Remainder of penial bulb inside body appears diffuse, to 340 Μm high. Prostate-free portion of the atrium 1460–1630 Μm long by 60–72 Μm wide, with muscle layer to 12 Μm thick ectally, 4 Μm entally; atrial duct narrows to 30 Μm within the penis, extending another 500 Μm. Ental, prostate-covered part of atrium 1380 Μm long by 60–79 Μm wide, with 2–3 Μm muscle layer and granular epithelium 7–8 Μm thick; prostate glands 100–120 Μm long and densely packed. Posterior vas deferens 23–28 Μm wide, anterior vas 15–17 Μm wide in XI–XII.
Remarks. Rhynchelmis diespluviae is associated with the yakimorum complex within R. ( Sutroa ) (Appendix 3). Spermathecal characters include the dense ectal bulbs, inconspicuous ducts, pyriform ampullae, and paired, branched diverticula. The elongate atria, with vasa deferentia entering within the ental, prostate-bearing portion, are typical for the subgenus. Based on the positions of the genital pores, the elongate spermathecal diverticula, and the conical penes, R. diespluviae appears closest to R. utahensis . However, the long, straight penes have terminal pores, and protrude into the sac even when the penial structure is completely retracted. In contrast, R. utahensis penes are sharply curved when everted, and they are short, with subterminal pores when retracted (Figs 36–41 in Fend & Brinkhurst 2000). The penial bulbs are smaller in R. diespluviae , the atria enter the penial bulb anterodorsally (rather than laterally), and the atrial musculature is thicker than in typical R. utahensis . The narrower spermathecal ampullae also distinguish the new species from R. utahensis .
The penes most closely resemble those of R. rostrata ( Fig. 1 View FIGURE 1. R B, C), but the male pores are more widely separated and the spermathecae are paired, with only two diverticula. Two R. diespluviae specimens had only one spermatheca, but these retained the ventrolateral position of normal specimens, in contrast to the consistently median spermatheca of R. rostrata . The spermathecal pores are located near the anterior septum in VIII, rather than slightly anterior to the ventral chaetae. Vasa deferentia are relatively thick compared with most other R. ( Sutroa ) species (cf. Figs 11 and 43 in Fend & Brinkhurst 2000), except for R. aleutensis ( Fig. 2 View FIGURE 2. R B in Fend 2005); however, they do not appear glandular, as in R. ( Rhynchelmis ) (cf. Fig. 2 View FIGURE 2. R G–H in Fend & Brinkhurst 2010).
Blood vessels were obscure in most specimens, but where visible, they appeared variable. There was only one obvious lateral pair in some segments, but other segments clearly had two. In addition, branching of these vessels appeared to be minimal compared with most R. ( Sutroa ) species (Appendix 3). Specimens from Orogrande Creek had smaller spermathecae and penial structures, and much weaker atrial musculature ( Fig. 8 View FIGURE 8. R G–H), but the basic form of these structures resembled those from the type locality, and differed from other species known from the region ( Fend & Brinkhurst 2000). The single specimen from Hurst Creek, Washington represents a considerable range extension, and the everted condition of its penial bulbs makes comparison with the Idaho material difficult. Nevertheless, the entire structure conforms to the expected everted state of the Idaho material, and differs from other R. ( Sutroa ) species.
Distribution. Most Rhynchelmis diespluviae records are from northern Idaho, mostly in tributaries of the Clearwater River. The distribution overlaps with other members of the R. yakimorum complex, particularly R. monsserratus , although the latter has been found mostly in the adjacent Salmon River drainage ( Fend & Brinkhurst 2000). The R. diespluviae distribution appears quite disjunct from that of the morphologically similar R. utahensis in southern Idaho and northern Utah. The latter appears restricted to upland rivers and springs in the Great Salt Lake drainage, with genetically distinct populations in nearby isolated springs associated with the Snake River ( Zhou et al. 2010). The widespread R. rostrata is not known from the area. Two R. (Rhynchelmoides) species, R. elrodi and R. saxosa Fend & Brinkhurst, 2000 , are common in the region. The Washington collection appears biogeographically disjunct from the Idaho material, and the “Clearwater River”, to which Hurst Creek is tributary, is a Pacific Coastal drainage not associated with the Idaho Clearwater River in the Columbia River drainage.


| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |