
Pyura carmanae, Rocha & Counts, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4564.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:FDADF9CA-379C-4980-8AE4-D47F58537E3F |
|
DOI |
https://doi.org/10.5281/zenodo.5930324 |
|
persistent identifier |
https://treatment.plazi.org/id/E0CB2AD6-478D-418C-A8A0-F05383594564 |
|
taxon LSID |
lsid:zoobank.org:act:E0CB2AD6-478D-418C-A8A0-F05383594564 |
|
treatment provided by |
Plazi |
|
scientific name |
Pyura carmanae |
| status |
sp. nov. |
Pyura carmanae View in CoL sp. nov.
( Figures 1 View FIGURE 1 , 2 View FIGURE 2 )
urn:lsid:zoobank.org:act:E0CB2AD6-478D-418C-A8A0-F05383594564
Pyura View in CoL sp. aff. vittata: Tokioka, 1972: 399 View in CoL .
Materials examined: Holotype: MZUSP553 View Materials one individual, Isla Canales de Tierra , Shark Point, 7°44'18”N 81°34'46.8”W, leg. R. M. Rocha, 10.01.2009. GoogleMaps
Paratype: DZUP PYU-127 seven individuals, Isla Canales , Shark Point, 7°44'18”N 81°34'46.8”W, leg. R. M. Rocha, 10.01.2009 GoogleMaps .
Etymology. The name of the species is a homage to Mary Carman from Woods Hole Oceanographic Institution who coordinated the expedition that found the species and for her important work concerning the study and management of invasive ascidians.
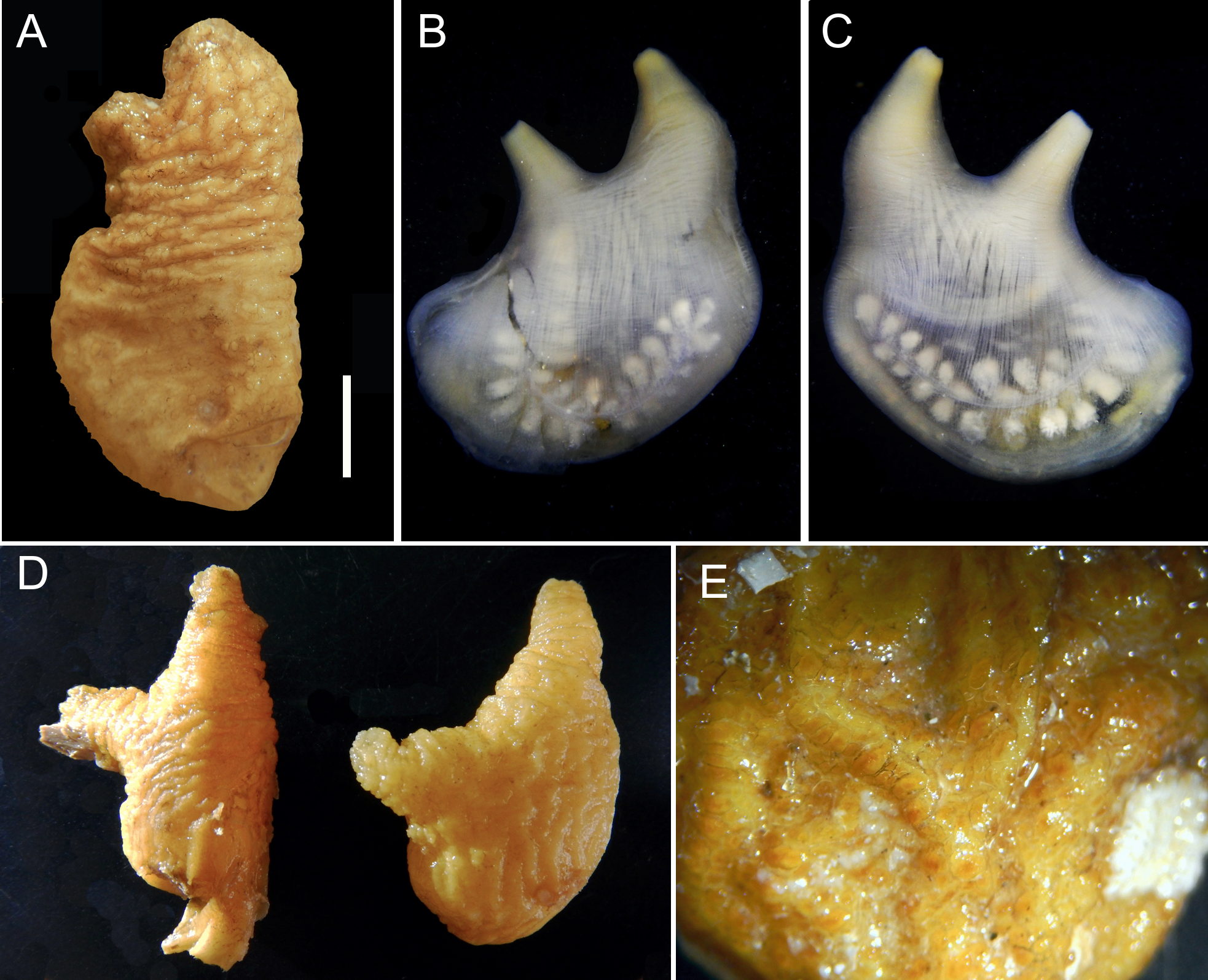
Description. Living specimens are yellow and wrinkled ( Fig. 1A, D View FIGURE 1 ); often found under rocks attached by the posterior region of the left side. The animals are usually 3–4 cm long but can reach 5.5 cm by 2 cm and are very flat (the holotype 3.5 cm long and 2.5 cm in diameter). The siphons are bumpy, apical and can be close together with little space separating the two or a little separated with the atrial siphon more lateral. The oral siphon is typically longer than the atrial siphon, but length varies among individuals ( Fig. 1D View FIGURE 1 ), and none of the siphons have spinules on the border or on their internal lining. After fixation, animals still retain the yellow coloration, but the tunic is white inside and presents many bumps and horizontal wrinkles; notably in the anterior half of the animal. The rough tunic is relatively thick (1–3 mm), and can show a tessellate pattern in the posterior half ( Fig. 1E View FIGURE 1 ). The tunic is usually clean of encrustations, but some bryozoans and entoprocts sparsely cover the surface.
The body wall coloration varies. In fresh specimens, red coloration can be seen on the siphons. After fixation, it is often white or a faint yellow. The musculature of the siphons is strong. Circular muscles are heavily dense around the siphons, making them contracted. Longitudinal fibers placed tightly together radiate from the siphons down the body resembling a continuous sheet of muscles, however, the U-shaped right gonad is often seen through the body wall ( Fig. 1B, C View FIGURE 1 ).
There are 22 laterally flattened oral tentacles in the holotype on a ring of strong circular muscles embedded in the body wall, but the number ranges from 20 to 30 among samples ( Fig. 2C View FIGURE 2 ). Tentacles are in one or two size orders but some specimens the tentacles mainly show one size, with a few smaller in between. Tentacles are thin, have a larger base that tapers off towards the end, and mostly have second order ramifications only; however, third order ramifications were viewed in some samples. The prepharyngeal groove is made up of two equal size lamellae, very close to the line of tentacles, forming a shallow V around the dorsal tubercle, which can be completely hidden under the tentacles. The aperture of the dorsal tubercle is U-shaped. The dorsal lamina does not begin immediately after the dorsal tubercle; instead, a small distance separates the two. The dorsal lamina is composed of long languets; 80 in the holotype. The pharynx is attached to the body wall by slender vessels.
The pharynx is uncolored with six folds per side ( Fig. 2A View FIGURE 2 ). In some cases, the right side folds are higher than the left side, and the ventral folds are reduced. Although high, the folds are separated from each other and do not overlap. The holotype has 231 longitudinal vessels and the formula is (from right to left side):
E 3 (8) 2 (13) 2 (16) 3 (19) 2 (20) 4 (21) 4 DL 1 (24) 3 (18) 0 (20) 3 (16) 3 (10) 8 (7) 6 E
While another smaller (2.5 cm) exemplar had the following formula (177 vessels in total):
E 3 (6) 4 (9) 4 (13) 3 (12) 3 (11) 3 (19) 5 DL 1 (16) 2 (14) 3 (15) 3 (14) 3 (10) 2 (6) 3 E
Parastigmatic vessels are present in many regions of the pharynx. There are four to six stigmata per mesh in between folds, and four or five on folds ( Fig. 2F View FIGURE 2 ).
Dense, irregular shaped endocarps are present on both sides of the animal. Those on the right are attached to the body wall dorsal and anterior to the right gonad, but absent between the right gonad and endostyle. Those on the left are attached to the body wall dorsal and anterior to the gut loop and there are also a few endocarps inside the primary intestinal loop ( Fig. 2B, E View FIGURE 2 ). Endocarps usually not present on the gut although in some specimens one or two small endocarps are on the descending intestinal wall. Endocarps are also present on the most anterior gonadal lobes. On several specimens, a few endocarps anteriorly placed looked to be storing an unknown substance.
The primary intestinal loop is large, with the ascending intestine running along the endostyle, but does not extending up to the prepharyngeal groove, although it surpass the level of the anus; the descending intestine is short and forms a shallow secondary loop with the rectum extending all the way inside the atrial siphon in specimens with siphons closer to each other ( Fig. 2B View FIGURE 2 ). In specimens with siphons a little bit more spaced, the primary loop is more narrow and the descending intestine longer, forming a more posterior secondary loop ( Fig. 2E View FIGURE 2 ). The intestine is isodiametric, transparent, and there can be a constriction before the anus, which has a smooth rim, sometimes enrolled outwards ( Fig. 2G View FIGURE 2 ). The digestive gland lies along the esophagus and stomach, with many small portions followed by a large and elongated mass of tubular projections, often yellow or green, with one connection with the stomach wall ( Fig. 2D View FIGURE 2 ).
The left gonad is spread inside the primary intestinal loop, the lobes have tubular connections with the main duct and are separated from each other ( Fig. 2B View FIGURE 2 ). The left gonad has 18–35 lobes. The lobes vary in size, are irregularly shaped, although tend to be oval. The right gonad is ventral and forms a deep U in the posterior half of the animal, with 28–35 lobes ( Fig. 2B View FIGURE 2 ). Oviduct and sperm duct are slightly lobed and equally long, opening right at the base of the atrial siphon ( Fig. 2G View FIGURE 2 ).
Remarks. Tokioka (1972) described 12 individuals of this species from Playas del Coco, Costa Rica as Pyura aff. vittata and he argued that character variation in P. vittata was not enough known at that time for the separation of that population as a new species. Indeed, the right gonad of P. carmanae has the same shape as in P. vittata , but the dilated rectum, presence of endocarps on the intestine and gonads, larger number of longitudinal vessels and presence of spinules in the siphons in P. vittata distinguishes this species from P. carmanae . Further, in Central America, P. vittata is only found in the Atlantic coast both on mangrove prop roots, coral reefs and artificial substrates ( Rocha et al. 2005; Carman et al. 2011), while P. carmanae is only found under rocks in the Pacific coast. Thus, the known geographical distribution of this species extends north at least until Costa Rica. Pyura discrepans ( Sluiter, 1898) known from Colombia and Martinique is similar to P. carmanae . Main differences are the brownish-red tunic, irregular stigmata in the pharynx, presence of abundant endocarps on the descending intestine in the holotype but not in the specimen from Martinique and gonadal lobes not well attached to the body wall in P. discrepans (see Sluiter 1898, Monniot 1983, Monniot 2018). Pyura ocellata Monniot, 2016 recently described from French Guiana is also similar to P. carmanae . Main differences are the smaller size, short siphons, smaller number of stigmata per mesh, larger digestive track, larger number of gonadal lobes in P. ocellata (see Monniot, 2016). Among the samples in the Smithsonian Museum collection, USMN 0 14621 and USMN 0 14666 collected from Venado Beach, Panama and identified as P. vittata , actually belong in this species.

| DZUP |
Universidade Federal do Parana, Colecao de Entomologia Pe. Jesus Santiago Moure |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Tunicata |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pyura carmanae
| Rocha, Rosana M. & Counts, Bailey Keegan 2019 |
Pyura
| Tokioka, T. 1972: 399 |