
Pyrgoma spurtruncata, Kim & Chan & Corbari & Rodriguez Moreno & Achituv & Kim, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4695.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:3007F762-721A-477B-94C9-AC0A24E2ACB8 |
|
persistent identifier |
https://treatment.plazi.org/id/9EA455E4-3168-4274-8B41-66AA11643C35 |
|
taxon LSID |
lsid:zoobank.org:act:9EA455E4-3168-4274-8B41-66AA11643C35 |
|
treatment provided by |
Plazi |
|
scientific name |
Pyrgoma spurtruncata |
| status |
sp. nov. |
Pyrgoma spurtruncata sp. nov.
( Figs 2−10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Type material. Holotype: MNHN-IU-2014-23572, New Caledonia, Norfolk Ridge , Jumeau Ouest seamount, on Madrepora oculata Linnaeus, 1758 ; SMIB 3 cruise (Stn DW 14, 23° 40’ 30’’ S, 167° 57’42’’ E, 22 May 1987, 246 m depth). GoogleMaps
Paratypes: MNHN-IU-2014-23573, data same as for holotype.
MNHN-IU-2014-23574, New Caledonia, Norfolk Ridge, Banc Kaimon, Maru seamount, on Madrepora oculata ; SMIB 4 cruise (Stn DW41, 24° 44’ 18’’ S, 168° 08’ 42’’ E; 8 March 1989; 230-235 m depth).
MNHN-IU-2014-5596, MNHN-IU-2014-5597, MNHN-IU-2016-9936, New Caledonia, Norfolk Ridge, Banc Antigonia, on Madrepora oculata ; SMIB 4 cruise (Stn DW55, 23° 20’ 48’’ S, 168° 4’ 48’’ E; 9 March 1985; 215- 260 m depth). MNHN-IU-2014-23575, New Caledonia, Norfolk Ridge, on Madrepora oculata ; BIOCAL cruise (Stn CP110, 22° 13’ 19’’ S, 167° 09’ 16’’ E; 9 September 1985; 275-320 m depth).
Diagnosis. Scutum triangular, transversally elongated, occludent and basal margins longer than rostral margin. Adductor plate thick; rostral tooth blunt, wide. Tergum upper part triangular, long narrow spur with moderately broad external furrow; without crests for depressor muscle.
Description (mainly based on holotype, except where stated). Shell ovate ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 ) plates totally fused, flat or low conical, white to pale yellow in preserved specimens ( Fig. 2 View FIGURE 2 ). External surface with 21 low, broad ridges radiating from orifice to shell circumference ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 ). Paratypes and other specimens with number of ridges ranging between 19−21 ( Fig. 2C View FIGURE 2 ). Internal surface of wall smooth, with 21 ribs at rim of shell ( Fig. 3A View FIGURE 3 ); sheath 1/3 height of shell, measured at rostral end ( Fig. 3A, C View FIGURE 3 ), surface with a number of growth rings ( Fig. 3C View FIGURE 3 ). Orifice ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 ) ovate, ~ 1/3 of basal rostro-carinal diameter. Basis of shell ( Fig. 3A View FIGURE 3 ) moderately deep, cup shaped ( Fig. 2B, D, E, F View FIGURE 2 ). Top region of basis, at junction between shell and basis, with simple sulci fitting short, longitudinal ribs developed from internal surface of shell ( Fig. 4A, E, F View FIGURE 4 ). In paratypes MNHN-IU-2014-23573 and MNHN-IU-2014-5597, top of basis with short sections of ladder-like, longitudinal perforations ( Fig. 4 View FIGURE 4 B-D, G-I). In cross-section view ( Fig. 4C, D View FIGURE 4 ), basis developing into extensive ladder-like, longitudinal perforations ( Fig. 5 View FIGURE 5 ); between ladder-like longitudinal perforations, single-row of deep grooves located between ladder-like longitudinal ridges, groove widening at shell rims ( Fig. 5C, D View FIGURE 5 ).
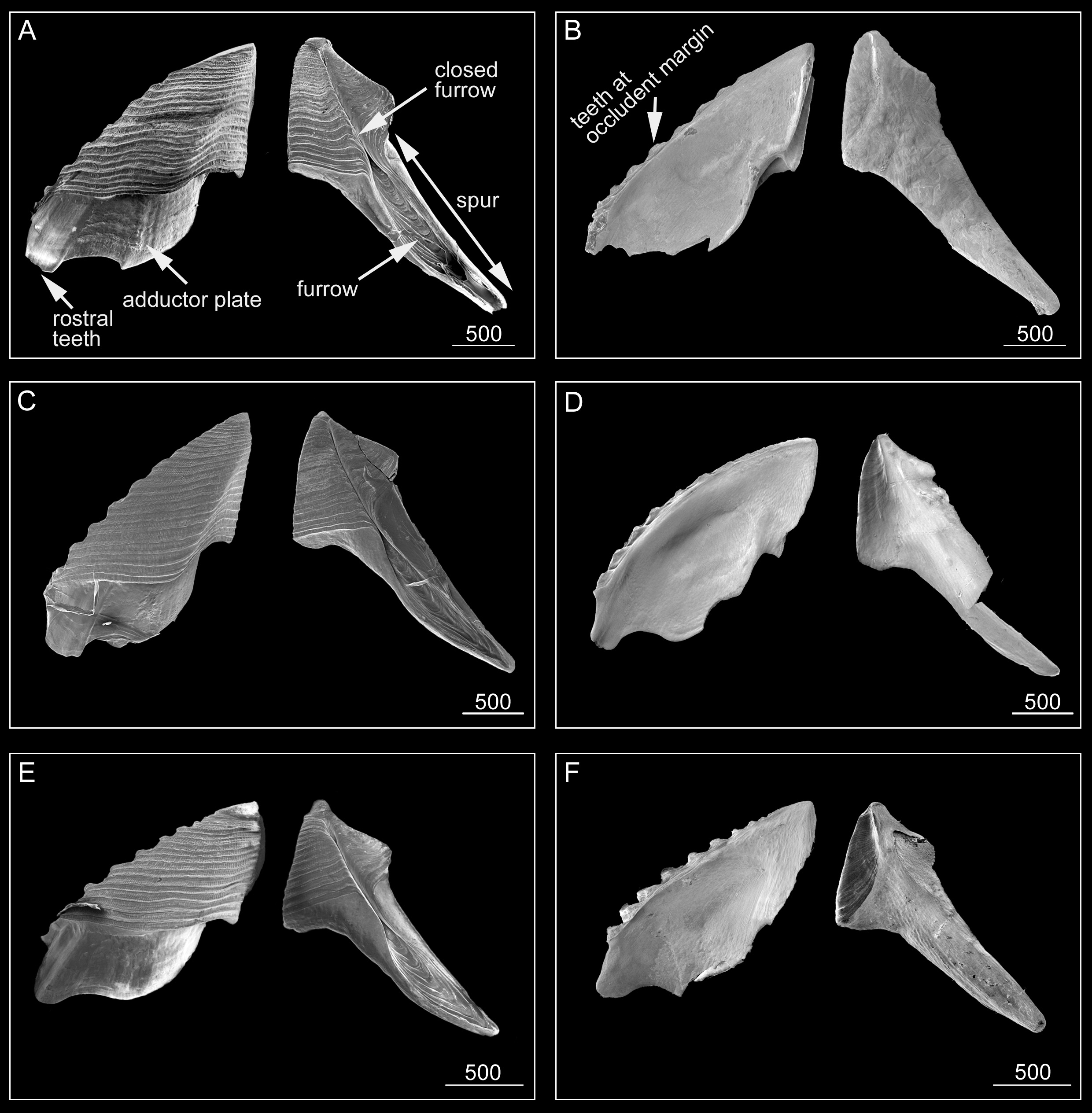
Scutum ( Fig. 6 View FIGURE 6 ) triangular, height similar to width, several small teeth along ventral surface ( Fig. 6B, D, F View FIGURE 6 ) of occludent margin. Tergal margin straight, less than 1/2 length of occludent margin. External view with visible adductor plate, deeply projecting below basal margin, reaching 1/3 height of scutum ( Fig. 6A, C, E View FIGURE 6 ). Adductor plate with blunt, thick, short rostral tooth ( Fig. 6A, C, E View FIGURE 6 ), height of tooth less than 1/3 height of adductor plate ( Fig. 6A, B View FIGURE 6 ). External surface with slightly sinuous striations, striations bearing rows of small pores. Internal surface with oval-shaped adductor muscle scar ( Fig. 6B, D, F View FIGURE 6 ). Scutum of paratypes, consistent with holotype ( Fig. 6A, C, E View FIGURE 6 ).
Tergum ( Fig. 6 View FIGURE 6 ) moderately broad, upper part triangular, higher than wide, height to width ratio 3:1, apical angle 68°. Distinct spur with furrow reaching 3/5 height of tergum; medial furrow fused at mid length of tergum, distal end of spur truncated ( Fig. 6 View FIGURE 6 ). External surface with slightly sinuous growth lines; basal margin not obvious due to spur ( Fig. 6A, C, E View FIGURE 6 ). Internal surface without crests for depressor muscle ( Fig. 6 View FIGURE 6 , B, D, F).
Maxilla ( Fig. 7A View FIGURE 7 ) bilobed, with dense, serrulate setae on margins of both lobes. Mandibular palp ( Fig. 7B View FIGURE 7 ) ovate, bearing simple setae on distal end and outer and inner margins. Maxillule ( Fig. 7C View FIGURE 7 ) with small notch, 2 large setae above notch, row of 8 developed setae below notch; outer and inner margins with long, simple setae. Mandible ( Fig. 7 View FIGURE 7 D−F) with 5 teeth excluding inferior angle; second to fourth teeth bifid; first 3 teeth occupying 3/4 length of cutting edge; fifth tooth blunt, close to inferior angle. Inferior angle with few large setae. Lateral side, lower margin and cutting edge of mandible bearing simple setae. Labrum ( Fig. 7G, H View FIGURE 7 ) with deep, V-shaped notch separating crests; 2 small teeth on each crest.
Cirrus I ( Fig. 8 View FIGURE 8 A−C) with unequal rami; anterior ramus long, slender, 12 segmented, bearing serrulate setae; posterior ramus short, stout, 6 segmented, bearing plumose setae on first segment ( Fig. 8B View FIGURE 8 ). Cirrus II ( Fig. 8D, E, G View FIGURE 8 ) with un-equal rami; anterior ramus 7 segmented, bearing simple setae, posterior ramus 5 segmented, first segment bearing plumose setae. Cirrus III ( Fig. 8F, H, I View FIGURE 8 ) anterior ramus 9 segmented, slightly longer than posterior ramus (7 segmented), first segment of posterior ramus bearing plumose setae. Cirri IV−VI ( Fig. 9 View FIGURE 9 ) long, slender, rami of equal size. Cirrus IV ( Fig. 9A View FIGURE 9 ) with anterior ramus 13 segmented, posterior ramus 15 segmented; cirrus V ( Fig. 9C View FIGURE 9 ) with anterior ramus 16 segmented, posterior ramus 17 segmented, cirrus VI ( Fig. 9E View FIGURE 9 ) with anterior ramus 16 segmented, posterior ramus 14 segmented. Intermediate segments of cirri IV−VI ( Fig. 9B, D, F View FIGURE 9 ) with 3 pairs of simple setae, distal two pairs longest, proximal pair shortest.
Penis ( Fig. 9G, H View FIGURE 9 ) long (~1.2 times length of cirrus VI), annulated, basidorsal point acute, setae simple; apex of penis bearing short, simple setae ( Fig. 9I View FIGURE 9 ).
Etymology. Compared to P. japonica and P. cancallatum , P. spurtruncata sp. nov. has a wider, shorter tergal spur, and a blunt rostral tooth of the scutum. The specific name spurtruncata refers to the wide, short spur of the tergum (the Latin word truncata means short).
Distribution. Presently recorded only from the deep-waters of New Caledonia and only from the coral Madrepora oculata in the family Oculinidae .
Remarks. In their revision of Pyrgoma, Ross and Newman (2002) divided Pyrgoma into two groups, based on the presence/absence of extended muscles crests of the tergum. Pyrgoma cancellatum ( Fig. 10A, B, E View FIGURE 10 ), P. japonica ( Fig. 10C, D View FIGURE 10 ) and P. kuri (see Fig. 2 View FIGURE 2 for SEM plate of scutum and tergum in Ross & Newman, 2000) lack tergal muscle crests. In contrast, P. projectum ( Fig. 10F View FIGURE 10 ) and P. sinica ( Fig. 10G View FIGURE 10 ) have extended tergal muscle crests on the tergum. Pyrgoma spurtruncata sp. nov. is morphologically similar to P. cancellatum , P. japonica and P. kuri in sharing an absence of tergal muscle crests. Pyrgoma spurtruncata sp. nov., however, can be distinguished from P. cancellatum by the shape of the medial furrow on the external surface of the tergum ( Fig. 10A, B, E View FIGURE 10 ). In P. cancellatum ( Fig. 10B View FIGURE 10 ), the medial furrow of the tergum is shallow and fully open. In P. spurtruncata sp. nov., the medial furrow is deeper and closed along half the length of the tergum. Pyrgoma spurtruncata sp. nov. is similar to P. japonica in that the rostral teeth of both species are distinct and blunt, and the external furrow of the tergum is closed. Compared to P. japonica , the tergal spur of P. spurtruncata sp. nov. is much wider, and the ratio of length to width of the spur is 5.5:1 (compared to the slender and narrow tergal spur in P. japonica with a ratio of 9:1) ( Fig. 10D View FIGURE 10 ). Similar to P. spurtruncata sp. nov., P. kuri has a truncated distal end of the spur of the tergum, but in P. kuri the tergal furrow is open (see Figure 2 View FIGURE 2 in Ross & Newman, 2000), whilst P. spurtruncata sp. nov. has a closed tergal furrow.
Most of the coral-inhabiting barnacles of the family Pyrgomatidae Gray, 1825 (e.g., Cantellius , Galkinius ) have a solid basis. In Pyrgoma , the basis has perforations or arches that are believed to provide chemical mediations between the coral hosts and the barnacles ( Ross & Newman, 2000). Ross and Newman (2000) examined the basis of P. kuri and found that the initial stage is ladder-like perforations, arranged longitudinally, that turn into arches when approaching the top of the shells. Additionally, the basis of P. cancellatum becomes arch-like as the basis develops. Conversely, P. spurtruncata sp. nov. shows some variations in the basis. The top of the basis (junction between shell and basis) in the holotype does not contain extensive longitudinal perforations and is mostly solid in nature ( Fig. 4A, E, F View FIGURE 4 ) but, in some specimens, there are some ladder-like, longitudinal perforations extending from the deeper to the upper part of the basis ( Figs. 4 View FIGURE 4 B-D, 4G-I, 5). However, no arches were seen on the joint with the basis. It is probable that P. spurtruncata sp. nov. does not develop arch-like perforations.
Molecular analysis. A total of nine sequences of COI and 12s rDNA from nine specimens (including four Pyrgoma spurtruncata sp. nov., one P. japonica and four P. cancellatum ) were amplified and compared to Pyrgoma sequences retrieved from GenBank. The sequences from the present study are deposited in GenBank ( Table 1 View TABLE 1 ). The concatenated alignment consisted of 452 nucleotides of 12S rDNA and 563 of COI.
Molecular identification using both mitochondrial 12S rDNA and COI gene sequences complemented the morpholoical identifications. Sequence similarity (K2P distance, Table 2 View TABLE 2 ) and the phylogenetic analyses (Maximum Likelihood, Fig. 11 View FIGURE 11 ) confirmed the identity of four P. spurtruncata sp. nov. specimens; the phylograms based on the two markers showed high similarities ( Fig. 11A, B View FIGURE 11 ). The phylogeny of the concatenated data set was similar to the phylogenetic trees derived from individual genes ( Fig. 11C View FIGURE 11 ).
In pairwise comparisons, the K2P distance among the sequences ranged from 0.1% to 18.8% ( Table 2 View TABLE 2 ). The mean intraspecific divergence of P. spurtruncata sp. nov. was only 0.1%. In contrast, the interspecific divergence (based on different species name from GenBank) of other Pyrgoma species ranged from 2.4 % to 15.6%. The low divergence value of 2.4% is between Pyrgoma sp. 1 ( Malay & Michonneau, 2014) and P. japonica , which located in the same clade in the molecular phylogenetic tree ( Fig. 11 View FIGURE 11 ), suggesting Pyrgoma sp. 1 from Malay and Michonneau, 2014 is P. japonica . Among the Pyrgoma species, P. spurtruncata sp. nov. shows a sister group relationship with P. japonica and the pairwise distance was 11.9% between two species.









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |