
Psychotria davisiana C.M. Taylor, 2020
|
publication ID |
https://doi.org/ 10.15553/c2020v752a1 |
|
DOI |
https://doi.org/10.5281/zenodo.6335852 |
|
persistent identifier |
https://treatment.plazi.org/id/03DFC37D-FFDC-FFD1-FFEE-F92C827390C2 |
|
treatment provided by |
Carolina |
|
scientific name |
Psychotria davisiana C.M. Taylor |
| status |
sp. nov. |
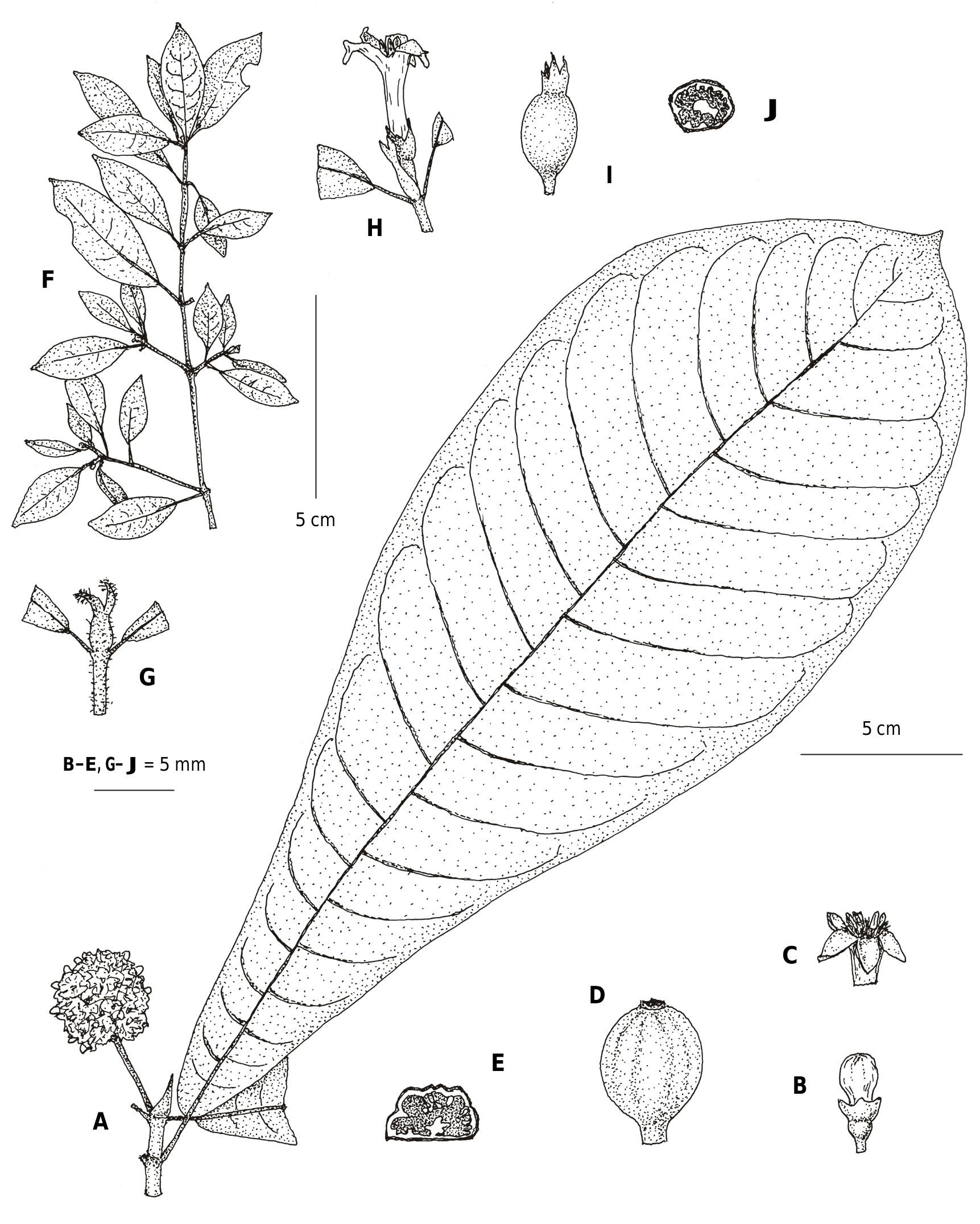
Psychotria davisiana C.M. Taylor View in CoL , sp. nov. ( Fig. 2F–J View Fig , 4 View Fig ).
Holotypus: MADAGASCAR. Reg. Analamanga [Prov. Antananarivo ]: Anjozorobe , Betatao , forêt de Vohitralongo , rte Anjozorobe-Ambatondrazaka , à 30 km au NE de Betatao, 18°04'07"S 47°58'41"E, 950–1093 m, 8.III.2000, Randrianaivo et al. 511 ( MO-6071774 !; GoogleMaps isotype: BR, G [ G00341940 ]!, K, MO-6146891 !, P, TAN) GoogleMaps .
Psychotria davisiana C.M. Taylor is distinguished from P. haplantha Bremek. by its larger leaf blades with acarodomatia, often shortly pedicellate flowers, longer calyx limbs, longer corollas, and endosperm that is at least shallowly ruminate on both abaxial and adaxial surfaces.
Shrubs and small trees, collected in flower and fruit variously at 1–4 m tall, much-branched, with well developed main stems bearing numerous branched lateral stems; stems weakly flattened when young becoming subterete, densely hirtellous to hispidulous with trichomes 0.2–0.5 mm, pubescence deciduous with age. Leaves opposite; petiole 0.1–0.5 cm, glabrous; blade oblanceolate to elliptic, 0.6–4.5 × 0.5–2 cm, at base cuneate to acute, at apex obtuse to acute or weakly acuminate with tip to 2 mm, drying papyraceous to chartaceous, on both surfaces glabrous; secondary veins 4 to 10 pairs, looping to interconnect near margins, usually with regularly developed small domatia comprising a pit and/or a tuft of strigose trichomes 0.1–0.3 mm, usually with 1 intersecondary vein present between pairs of secondary veins, adaxially costa thickened to prominent, secondary veins plane to thickened, and remaining venation plane, abaxially costa prominent, secondary and tertiary veins plane to prominulous, and remaining venation not visible. Stipules interpetiolar, caducous, abaxially and marginally densely hispidulous to hirtellous with trichomes 0.2–0.5 mm, adaxially glabrous in upper part and densely strigillose in lower part with trichomes 0.1–0.2 mm, oblong to pandurate in outline, 1.2–7 mm, lobed for 1/4–1/2 their length, basal portion (i.e., below lobes) ligulate to triangular or ovate, lobes lanceolate to narrowly triangular or linear, acute, erect when young then spreading laterally with age. Inflorescences terminal, reduced from a cyme to 1-flowered or shortly cymose and 2- or 3-flowered, enclosed by 1 pair of persistent stipules; bracts apparently reduced; pedicels 0.1–1 mm. Flowers variously subsessile to pedicellate, 5-merous, distylous; hypanthium obconic to cylindrical, c. 1 mm, glabrous to densely hirtellous with trichomes 0.3–0.5 mm; calyx limb abaxially and adaxially glabrous, tubular portion 1–2 mm, lobes narrowly ligulate to narrowly triangular, 1–2.5 mm, acute to obtuse or rounded, marginally ciliate; corolla salverform, yellow, externally glabrous, tube cylindrical, 5– 8 mm, 1.2–1.5 mm diam. near middle, internally glabrous except densely pilosulous at stamen insertion in upper part of tube, lobes triangular, 2–2.5 mm, acute, adaxially plane (i.e., not galeate), abaxially with conical thickening 0.5–1 mm near tip; stamens in short-styled form with filaments 2–2.5 mm, inserted in upper part of corolla tube, anthers c. 1.2 mm, exserted, in long-styled form not seen; style in short-styled form c. 3 mm with stigmas c. 1 mm, included, in long-styled form c. 7.5 mm with stigmas c. 1 mm, exserted. Infructescences similar to inflorescences. Fruits subglobose, c. 5 mm diam., sparsely hirtellous to glabrous, red, apparently juicy; pyrenes 2, hemispherical, adaxially plane, abaxially smooth to weakly irregularly ridged or wrinkled; endosperm adaxially sparsely but deeply ruminate, abaxially sparsely shallowly ruminate.
Etymology. – The epithet of this species honors Dr. Aaron Davis, a Rubiaceae specialist who has documented and studied Malagasy Psychotria ( DAVIS et al., 2007) .
Habitat, distribution and phenology. – Psychotria davisiana has been collected in humid, usually dense evergreen forest at 950–1342 m in central eastern Madagascar, and reportedly is often locally common. It has been collected with flowers in February, March, May, and June, and with ripe fruits in July and October.
Conservation status. – Psychotria davisiana is known from 19 specimen collections representing 19 unique occurrences in humid, usually dense evergreen forest at 950–1342 m elevation. The EOO of the species is 2,705 km ², within the limits for “Endangered” under IUCN Red List Criterion B1 ; and the AOO is 44 km ², also within the limits for “Endangered” under Criterion B2 ( IUCN, 2012). One occurrence is within the protected Anjozorobe Angavo forest corridor, one of the last vestiges of natural forests in Madagascar central highlands. The other 18 occurrences are in unprotected areas, one to the north and 17 to the south of the protected area. Forested areas without formal protection in the vicinity of Anjozorobe Angavo are subject to degradation by small-scale slash and burn agriculture and fire to create habitat for cattle farming ( GOODMAN et al., 2018). The one occurrence to the north of Anjozorobe Angavo (at Vohitralongo Forest) constitutes one location (sensu IUCN, 2012) with respect to the known threats; the one within the protected area is a second location; the two isolated occurrences to the south constitute one location each; and the 15 collection sites from the Moramanga area, the site of a long-standing nickel mining operation, are a fifth location. Given the nature of three of the locations with general habitat degradation in unprotected sites and the remaining location in close proximity to on-going mining operations, causing a decline in AOO and number of mature individuals, the Red List status of P. davisiana is assessed as “Endangered” [EN B1ab(ii,iii,v)+2ab(ii,iii,v)].
Notes. – Psychotria davisiana is characterized by its stems with smooth internodes and hirtellous pubescence; relatively small, obovate, abaxially glabrous leaves with small crypt-type domatia; hirtellous, rather small, caducous stipules that are bilobed for 1/3–1/2 of their length; terminal, subsessile to shortly pedunculate, solitary to three flowers subtended by a pair of persistent stipules; rather well developed, deeply lobed calyx limbs; medium-sized yellow corollas with the lobes abaxially bearing a conical projection; medium-sized red fruits; pyrenes that are smooth abaxially; and endosperm that is ruminate both abaxially and adaxially. The specimens seen dried brownish green or dark brown, with the dried trichomes dark reddish brown. Psychotria davisiana has a distinctive branching arrangement, with leafless principal stems bearing smaller, ascending, branched stems that do bear leaves. The acarodomatia are relatively small, and only one or two are developed on some leaves so these can be overlooked. On flowering branches, most of the stems bear inflorescences but few flowers are open at any time. Few long-styled flowers have been seen, and these were not dissected in order to preserve them. The style length is presented here following BREMEKAMP (1963) ’s format and based on one flower, but it probably varies with corolla size to position the stigmas just above the top of the corolla tube. This is one of several Psychotria species with relatively small leaves and the flowers borne singly or in small groups at the stem apices; only five such species were treated by Bremekamp, but this habit and flower presentation are now known from more species of Psychotria as well as some very similar species of Gaertnera Lam. (MALCOMBER & TAYLOR, 2009) . Psychotria davisiana agrees with Bremekamp’s Mapouria Group VII, which is diagnosed in part by endosperm that is ruminate both adaxially and abaxially; the adaxial ruminations in P. davisiana are well developed, while the abaxial ones seen are small and can be overlooked.
Psychotria davisiana is similar to P. haplantha Bremek. , which can be separated by its branching pattern, with welldeveloped leafless stems bearing regularly spaced, short, unbranched, leaf-bearing stems; usually smaller leaf blades, 0.6– 1.4 × 0.4–0.7 cm, without domatia; subsessile flowers; shorter calyx limbs, 0.8– 1 mm long; shorter corollas, with the tube 1.8 –3 mm long; and endosperm that is entire or ruminate only adaxially. The fruits of P. haplantha were not seen by BREMEKAMP (1963), but are now documented: these are red at mautiry, ellipsoid to subglobose, and c. 5.5 mm in diameter, with two pyrenes that are smooth to weakly longitudinally ridged abaxially and endosperm weakly ruminate on the adaxial face (Kotozafy 1006, MO) (Bremekamp included P. haplantha in his Psychotria Group I, but he did not know the fruits are red so it does not agree with the characters of that group.) Psychotria davisiana keys to Bremekamp’s Mapouria Group VII, or if the abaxial seed ruminations are not noted to his Mapouria Group V; all of the species in both of those groups differ from P. davisiana in their branched inflorescences with more than three flowers.
Paratypi. – MADAGASCAR. Reg. Analamanga [Prov. Antananarivo ]: piste de Tsiazompaniry à Andapa , Andramasina , [19°16'00"S 47°59'00"E], VI.1961, Service Forestier 20291 ( MO, P) GoogleMaps . Reg. Alaotra-Mangoro [Prov. Toamasina ]: Moramanga , Andasibe , Berano , Ambotavy forest , 18°51'52"S 48°17'50"E, 1018 m, 3.III.2005, Antilahimena et al. 3569 ( MO, P, TAN n.v.) GoogleMaps ; Ambatovy, Sahaviana forest , 18°51'37"S 48°17'10"E, 976 m, 14.II.2007, Antilahimena et al. 5345 ( MO, TAN) GoogleMaps ; Ambatovy , Andranovery forest , 18°52'04"S 48°18'27"E, 22.X.2007, Antilahimena 5886 ( MO, P, TAN) GoogleMaps ; ibid. loco, 18°51'55"S 48°18'35"E, 993 m, 10.II.2008, Antilahimena 6011 ( MO, P, TAN) GoogleMaps ; ibid. loco, 18°52'12"S 48°18'34"E, 1003 m, 15.II.2008, Antilahimena 6055 ( MO, P, TAN) GoogleMaps ; Ambatovy, côté SE de Berano , 18°49'18"S 48°17'04"E, 1046 m, 13.X.2008, Miandrimanana 389 ( MO, TAN) GoogleMaps ; Ambatovy, côté Ampitambe , 18°49'17 "S 48°17 '09"E, 1011 m, 13.X.2008, Miandrimanana 398 ( MO, P, TAN) GoogleMaps ; ibid. loco, 18°51'49"S 48°18'39"E, 988 m, 10.II.2008, Rakotondrafara et al. 542 ( MO, TAN) GoogleMaps ; ibid. loco, 18°51'53"S 48°18'36"E, 14.II.2008, Rakotondrafara et al. 622 ( MO, P, TAN) GoogleMaps ; ibid. loco, 18°50'47"S 48°17'18"E, 1020 m, 4.III.2005, Rakotovao et al. 1486 ( MO, P, TAN) GoogleMaps ; ibid. loco, 18°49'34"S 48°17'14"E, 1058 m, 25.X.2011, Rasoazanany et al. 29 ( AVY, MO, P, TAN) GoogleMaps ; ibid. loco, 18°51'05"S 48°18'13"E, 1080 m, 15.X.2005, Razanatsoa et al. 530 ( K, MO, P, TAN) GoogleMaps ; Phelps-Dodge project site , c. 15 airkm NE of Moramanga , c. 11 km E of Antanambao, 18°50'16"S 48°21'14"E, 950 m, 4.III.1997, Rakotomalaza et al. 1231 ( MO, P, TEF) GoogleMaps ; Ambongabe, Sahamorandro , 18°39'30"S 47°57'59"E, 1342 m, 8.IV.2005, Rakotonirina et al. 240 ( MO, P, TAN) GoogleMaps ; Ambatovy, Ankosy , 18°48'41"S 48°17'45"E, 1071 m, 31.VII.2012, Ramahenina et al. 85 ( MO, P, TAN) GoogleMaps ; Andasibe, Berano, Ampanadiofanantsy forest , 18°51'46"S 48°16'31"E, 983 m, 27.X.2008. Randrianasolo et al. 983 ( MO, TAN) GoogleMaps ; massif de l’Ifody , rive droite du Mangoro , [18°55'S 48°03'E], 29.V.1966, Service Forestier 24803 ( MO, P) GoogleMaps .

| AVY |
AVY |
| TEF |
TEF |
| BR |
Embrapa Agrobiology Diazothrophic Microbial Culture Collection |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
| K |
Royal Botanic Gardens |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| TAN |
Parc de Tsimbazaza |
| MO |
Missouri Botanical Garden |
| TEF |
Centre National de la Recherche Appliquée au Developement Rural |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |