
Pseudatheta elegans Cameron, 1920
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5319.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:45CD8E29-3320-42E2-A8A8-3A36BC0F41CE |
|
DOI |
https://doi.org/10.5281/zenodo.8184832 |
|
persistent identifier |
https://treatment.plazi.org/id/090C879D-FFED-8E10-899A-FF4DA98FFB1F |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudatheta elegans Cameron, 1920 |
| status |
|
Pseudatheta elegans Cameron, 1920 View in CoL
( Figs. 1F View FIGURE 1 , 3 View FIGURE 3 , 7E View FIGURE 7 –G)
[Japanese name: Ko-hime-kinokotsuyakeshi-hanekakushi]
Pseudatheta elegans Cameron, 1920: 225 View in CoL (original description;
type locality: “Bukit Timah, woodlands, Mandai” [ Singapore]): Cameron, 1939: 226 (Northern India); Schülke & Smetana, 2015: 634 (catalogue).
Material examined. Type materials compared. Syntypes: 1 male, “Fungus // Mandai, / Singapore. / Dr. Cameron. // SYN- / TYPE [blue round label pinned by a curator] // Pseudatheta / elegans Cam. / M. E. Bacchus det 1978 / SYNTYPE // Pseudatheta / elegans CAM. / det. R. Pace 1984 / LECTOTYPUS // NHMUK015025222 About NHMUK ” ( BMNH) ; 1 male, “Fungus / (rotten) // Mandai, / Singapore. / Dr. Cameron. // SYN- / TYPE [blue round label pinned by a curator] // Pseudatheta / elegans Cam / M. E. Bacchus det 1978 / SYNTYPE // Pseudatheta / elegans CAM. / det. R. Pace 1984 / PARALECTOTYPUS // NHMUK015020370 About NHMUK ” ( BMNH) ; 1 male, “SYN- / TYPE [blue round label pinned by a curator] // Mandai , / Singapore. / Dr. Cameron // TYPE / Pseudatheta / elegans . / Dr. CAMERON. // Pseudatheta / elegans Cam. / M. E. Bacchus det 1978 / SYNTYPE // Pseudatheta / elegans CAM. / det. R. Pace 1984 / PARALECTOTYPUS / Dissection By? // NHMUK015020337 About NHMUK ” ( BMNH) : 1 male, “Fungus / (rotten) // Woodlands, / Singapore. / Dr. Cameron. // SYN- / TYPE [blue round label pinned by a curator] // Pseudatheta / elegans CAM. / det. R. Pace 1984 / PARALECTOTYPUS // Pseudatheta / elegans Cam. / M. E. Bacchus det 1978 / SYNTYPE / NHMUK015025223 About NHMUK ” ( BMNH) ; 1 female, “Wood / (rotten) // Bukit Timah, / Singapore. / Dr. Cameron. / SYN- / TYPE [blue round label pinned by a curator] // Pseudatheta / elegans Cam. / M. E. Bacchus det 1978 / SYNTYPE // Pseudatheta / elegans CAM. / det. R. Pace 1984 / PARALECTOTYPUS // NHMUK015025224 About NHMUK ” ( BMNH)
Additional materials studied. JAPAN: [Miyagi-ken]: 1 male, Yoshida, Taiwa-chô, 20 VIII 2009, J. Aoki leg. ( KUM) ; [Fukuoka-ken]: 1 male, Tonoue-yama, Moji-ku, Kitakyûshû-shi, 15 V 2022, J.-H. Park leg. ( KUM) ; 1 male, 1 female, Hiko-san, Soeda-machi, 24 VII 2020, T. Nozaki leg. ( KUM) ; 1 male, Ryôgenji, Munakata-shi, 15 V 2022, T. Hashizume leg. ( KUM); [Saga-ken]: 1 male, 2 females, Uki-dake, Nanayama, Karatsu-shi, 6 X 2022, N. Tsuji leg. ( KUM) ; [Nagasaki-ken]: Tsushima Is .: 1 male, 1 female, Kamiagatamachi Sagohigashisato, Tsushima-shi, 18 VI 2022, T. Hashizume leg. ( KUM); 1 male, Kamiagatamachi Sasuna, 19 VI 2022, T. Hashizume leg. ( KUM); Fukue-jima Is .: 1 female, Kishikumachi Nakadake, Gotô-shi, 11 X 2022, T. Hashizume leg. ( KUM) ; [Oita-ken]: 1 male, Takatoriya-yama, Minamitabaru, Ume, Saeki-shi, 14 XI 2020, S. Inoue leg. ( KUM) ; [Kagoshima-ken]: 1 female, Sata-misaki, 5 V 1985, Y. Takai leg. ( KUM); Yaku-shima Is .: 2 males, 1 female, Koseda, Yakushima-chô, 22 VII 2021, T. Hashizume leg. ( KUM); Amami-Ôshima Is .: 1 male, Sumiyôchô Nishinakama, Amami-shi, 27 VI 2021, T. Hashizume leg. ( KUM); 1 male, Shinmura, 11 IV 1971, M. Sakai leg. ( EUM); Tokuno-shima Is .: 1 female, Mikyo, 12 IV 1968, M. Tomokuni leg. ( EUM) ; [Okinawa-ken] Ishigaki-jima Is .: 1 male, Omoto-dake, Hirae, Ishigaki-shi, 24 III 2022, S. Inoue leg. ( KUM) .
Redescription. Measurements (n = 5): BL ≈ 1.49–1.94; FBL, 0.86–0.96; HL, 0.29–0.30; HW, 0.34–0.36; PL, 0.30–0.34; PW, 0.44–0.48; EL, 0.30–0.35; EW, 0.51–0.56.
Relative length of antennomeres I–XI (n = 1): 23: 25: 18: 11: 10: 11: 10: 10: 11: 11: 31. Ratio of length/width of antennomeres I–XI (n = 1): 1.88: 2.35: 2.10: 0.96: 0.77: 0.75: 0.96: 0.58: 0.63: 0.57: 1.46.
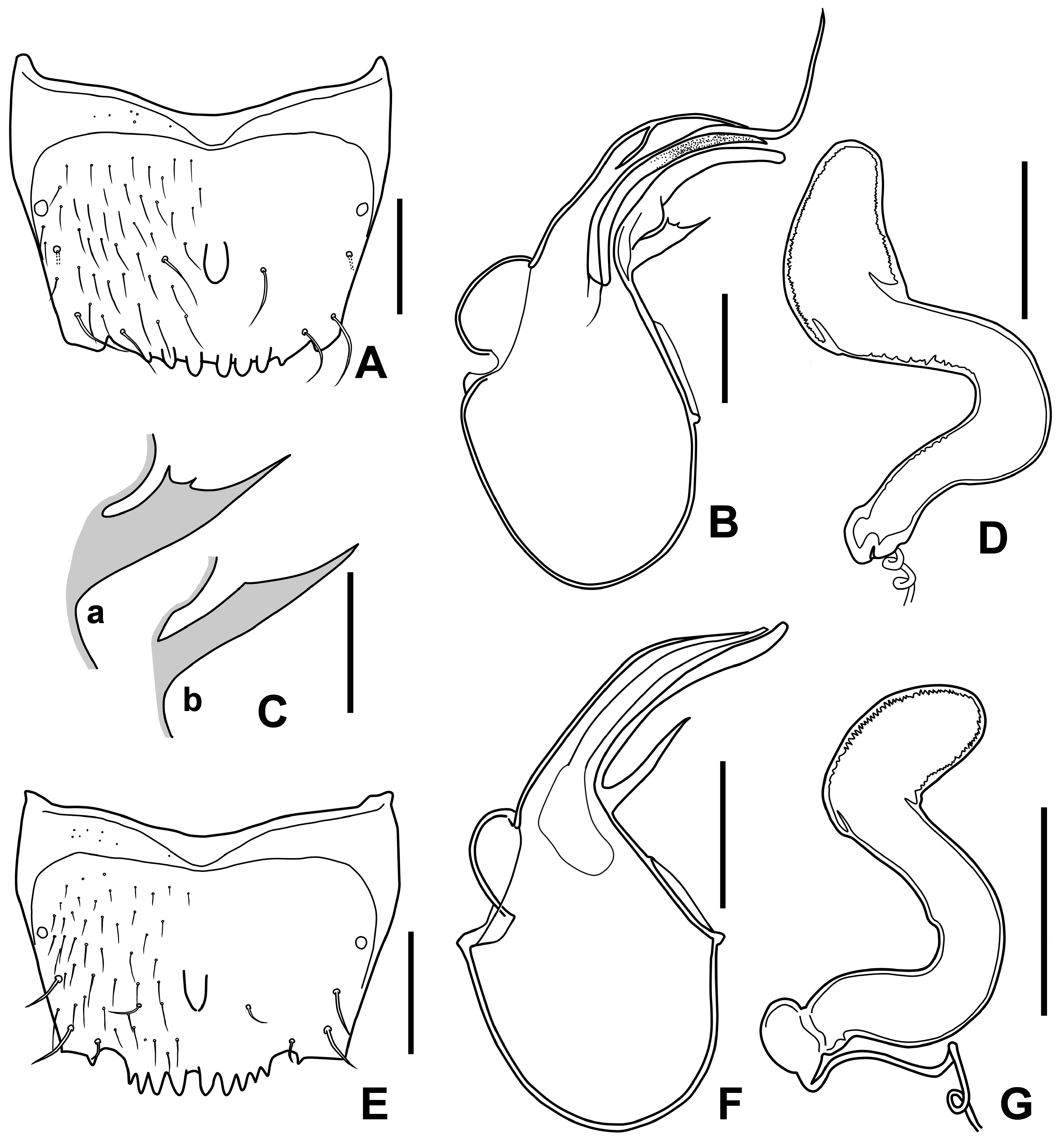
Body ( Fig. 1F View FIGURE 1 ) reddish brown; head darker; elytra dark brown except for the anterior 1/4–1/3; abdominal segments V–VI darker.
Head almost as long as wide, HW/HL:1.17–1.21; surface densely covered with setae.Antenna with antennomeres I–III and XI longer than wide, antennomeres IV–X wider than long; antennomeres I and II with rounded distal end, antennomeres III–X strongly angulated near distal end, antennomere XI oval.
Pronotum transverse, PW/PL: 1.30–1.46, PW/HW: 1.30–1.31; surface densely covered with setae, finely punctured, without microsculpture; posterior margin arcuate. Elytra wider than long, EW/EL: 1.54–1.71, EL/PL: 1.00–1.04, EW/PW: 1.17–1.29; surface densely covered with setae and finely punctured; posterior margin sinuate near posterolateral corners. Hind wings well developed; without flabellum. Mesoventrite without longitudinal carina; mesoventral process extended to slightly beyond middle of mesocoxal cavities, with pointed apex; metaventral process shorter than mesoventral process, with rounded apex; isthmus absent.
Abdomen slightly narrowed posteriad; surface densely covered with setae.
Male. Elytra with a pair of small tubercles at about posterior 1/3 on sutural margin. Tergite VII with a tubercle on anteromedian area. Tergite VIII ( Fig. 7E View FIGURE 7 ) with a tubercle on median area; eight to ten processes on posterior margin. Aedeagus as in Fig. 7F View FIGURE 7 ; median lobe with large basal bulb; apical process elongated, sinuated, apical half slightly widened; ventral process narrow, sinuated; flagellum moderately long.
Female. Spermatheca ( Fig. 7G View FIGURE 7 ) curved twice; distal portion nearly reniform; median portion elongate, curved; proximal portion round, wider than long; constriction between median and proximal portion distinct.
Distribution. Japan (Honshu, Kyushu, Tsushima Is., Gotô-rettô Isls. (Fukue-jima Is.), Yaku-shima Is., Amami-Ôshima Is., Tokuno-shima Is., Ishigaki-jima Is.)—new record; Singapore, India.
Remarks. We compared photographs of the habitus and the aedeagi of the type series with those of the specimens collected from Japan, and determined them to be the same species based on the similarity of dorsal habitus, the slightly sinuate apical process of the median lobe of aedeagus, and moderately long flagellum of the aedeagus. This species is similar in the shapes of the aedeagus and the spermatheca to Ps. tronqueti , described from Sri Lanka by Pace (1988), Ps. mendica , described from Nepal by Pace (1989), Ps. rougemonti , described from Sabah in East Malaysia by Pace (2014), Ps. borneensis , described from Sabah by Pace (2007), Ps. kinabaluensis , described from Sabah by Pace (2007), and Ps. pahangensis , described from Peninsular Malaysia by Pace (2012). These similar species, except for Ps. mendica , can be distinguished from Ps. elegans as follows:
in Ps. rougemonti , the ventral process of the median lobe of aedeagus is almost straight;
in Ps. borneensis the flagellum of the median lobe of aedeagus is longer, and the proximal portion of the spermatheca is slightly smaller;
in Ps. kinabaluensis , the ventral process of the median lobe of aedeagus is simply slightly curved dorsally, and the shape of the proximal portion of the spermatheca is different;
in Ps. tronqueti , the apical process of the median lobe of aedeagus is almost straight, and the proximal portion of the spermatheca is slightly smaller;
in Ps. pahangensis , the distal portion of the spermatheca is slightly broader and shorter. It is desirable to find males of Ps. pahangensis because the spermatheca is very similar to that of Ps. elegans .
Unfortunately, Ps. mendica cannot be distinguished from the Japanese Ps. elegans based on the information in the original description based on the female holotype ( Pace, 1989).
Some of species compared here are difficult to distinguish and we cannot refute the possibility that they are geographic or individual variation of this species. In fact, the aedeagus of the Pseudatheta sp. from Taiwan we examined is almost identical to those of specimens from Japan, but the distal portion of the spermatheca is slightly shorter. In this study, we were not able to examine the spermatheca of the type series of Ps. elegans and it is desirable to do so in the future.


| R |
Departamento de Geologia, Universidad de Chile |
| KUM |
Resource Management Support Center |
| V |
Royal British Columbia Museum - Herbarium |
| T |
Tavera, Department of Geology and Geophysics |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Aleocharinae |
|
Tribe |
Homalotini |
|
SubTribe |
Bolitocharina |
|
Genus |
Pseudatheta elegans Cameron, 1920
| Hashizume, Takuto, Yamamoto, Shûhei & Maruyama, Munetoshi 2023 |
Pseudatheta elegans
| Cameron, M. 1920: 225 |