
Pseudamnicola, Paulucci, 1878
|
publication ID |
https://doi.org/ 10.1080/00222933.2011.623358 |
|
persistent identifier |
https://treatment.plazi.org/id/03C9996E-FFE9-2C05-EB8F-8E7CFE55FE87 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudamnicola |
| status |
|
Pseudamnicola View in CoL ? (Corrosella) hydrobiopsis Boeters, 1999
Type locality
La Carmonilla spring in Loja, Granada, Spain, UTM: 30S 399062/4113653 .
Type material
The species is described based on the following type material: “ Holotype ( NNM 59145) and two paratypes ( NNM 59146) ex BOE 1399 ” ( Boeters 1999) .
Material examined
After exhaustively exploring the type locality and surrounding areas (according to the standard and Bou-Rouch methods used to collect the original shells) the species could not be found. The holotype, a dry shell, was borrowed from RMNH and examined and measured. One paratype was measured on the picture in the original description.
Description
Original description in Boeters (1999) ( Figures 1 View Figure 1 , 2 View Figure 2 ). Since no live specimens were found, the description below only includes shell features.
Shell shape turbinated, whitish as a result of erosion, height about 3.5 mm (Appendix Table 1), 5.5 shell whorls ( Figure 24A View Figure 24 ); in non-eroded spire whorls, longitudinal thin stripes can be observed; tip of shell, therefore protoconch, eroded ( Figure 24C View Figure 24 ); no deep suture between spire whorls and oblique; peristome frontal, complete, oval, with a pointed superior edge; thin outer lip and thicker inner lip which partially hides the umbilicus because of contact with body whorl; in lateral view, peristome is not straight since it is broken ( Figure 24B View Figure 24 ).
Remarks
This species was originally described based only on three shells found through “sondage Bou-Rouch” ( Boeters 1999). Ten years later this sampling method was repeated at the type locality, but no shells were found either there or at any surrounding locality. The description of the species therefore remains incomplete.
The type material and original description suggest that shell shape resembles species of Hydrobia Hartmann, 1821 more than Pseudamnicola . In fact, the name of the species refers to similar shell characters to those seen in Hydrobia ( Boeters 1999) . Given the lack of anatomical data and its similar shell features to the genus Hydrobia , it might be more appropriate to include it in this genus, in Moitessieria Bourguignat, 1863 , or a genus other than Pseudamnicola . However, until a live specimen is found, its generic placement remains uncertain.
One of the differentiating characters of so called P. (C.) hydrobiopsis is a turbinated shell and 5.5 spire whorls, differing from all the P. ( Corrosella ) species discovered to date. Pseudamnicola (C.) iruritai sp. nov. inhabits a spring near the type locality of P. (C.) hydrobiopsis , but the shell of P. (C.) iruritai sp. nov. differs in its proportions despite being of similar size. The body whorl and peristome of P. (C.) iruritai sp. nov. are larger and have a thicker inner lip completely hiding the umbilicus, whereas the umbilicus of P. (C.) hydrobiopsis is slit-like. Further, the spire whorls in P. (C.) hydrobiopsis are wider and bear more oblique suture.
Pseudamnicola (C.) luisi and P. (C.) marisolae sp. nov. are the closest species geographically, but are larger, more conical, with a larger aperture and umbilicus. Pseudamnicola (C.) falkneri and P. (C.) bareai sp. nov. inhabit the same province but further away, and are smaller, also more conical, with shorter and narrower spire whorls and a thicker inner lip in the peristome.
Statistical analysis
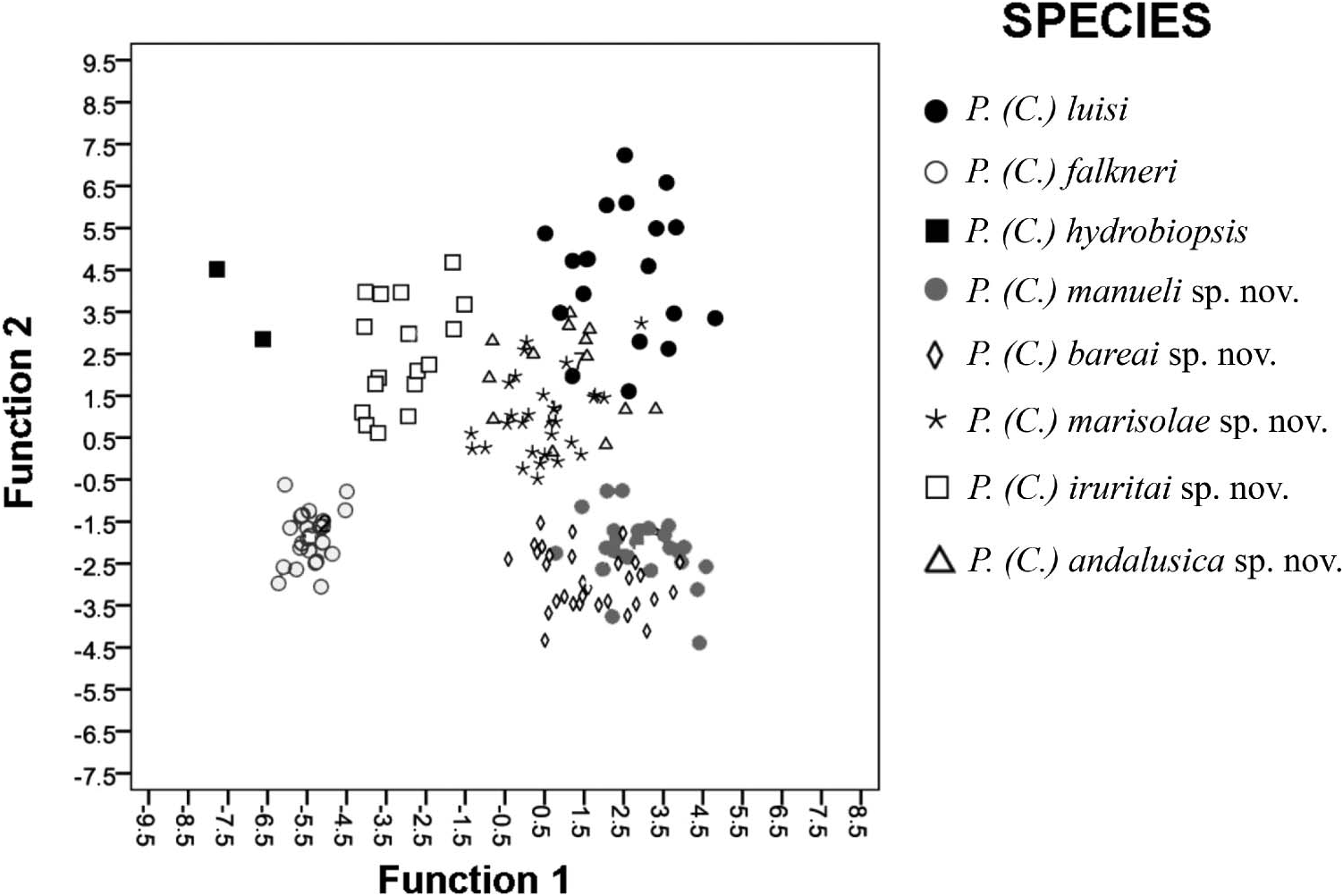
Discriminant function analysis identified seven highly significant discriminant functions (Wilk’s lambda = 0.00075; F(77, 864) = 26.045, P <0.001). The most discriminant variables included in these functions were: SW, AW, SL- LBW, SL and WPW. For the first function, which accounted for 49.6% of the explained variance with an eigenvalue of 9.27, the variables that most contributed (higher weight) to this variance were (in order): SW and AmW. For the second function, which accounted for 86.7% of the accumulated variance, the order was: SL-LBW, SL and WPW. All discriminant functions were highly significant (P <0.001). Of the 160 specimens examined, all P. (C.) falkneri , P.? (C) hydrobiopsis and P. (C.) iruritai sp. nov. were correctly classified (100%); 93% of P. (C.) marisolae sp. nov., 92% of P. (C.) ansalusica sp. nov. and P. (C.) manueli sp. nov., 90% of P. (C.) bareai sp. nov. and 84% of P. (C.) luisi individuals were also correctly classified. In the scatterplot ( Figure 25 View Figure 25 ) eight clusters can be observed. Two groups of most overlapping clusters emerged: the first one corresponding to two new species [ P. (C.) manueli sp. nov. and P. (C.) bareai sp. nov.], which share a similar shape and intermediate shell length and the lowest number of spire whorls. The second one comprised P. (C.) luisi , P. (C.) marisolae sp. nov. and P. (C.) andalusica sp. nov., which are the largest species of Corrosella .
Despite the shell shapes of the species examined being similar with the exception of P.? (C.) hydrobiopsis , some differences in size exist; P. (C.) falkneri and P. (C.) luisi had the most different shell sizes. In the scatterplot, populations appeared as two groups for the first function according to shell weight and aperture width. Pseudamnicola (C.) falkneri , P. (C.) iruritai sp. nov. and P.? (C.) hydrobiopsis appear in the same group because they have the narrowest shells, with a higher SL/SW index and narrower aperture. According to the second function, P. (C.) bareai sp. nov., P. (C.) manueli sp. nov. and P. (C.) falkneri are separated from the rest of the species because they show the shortest first spire whorl lengths (SL-LBW) (body whorl excluded).
DNA sequence data
A data matrix with 21 taxa and 658 characters for COI resulted in 14 different haplotypes. Of these 14 haplotypes, 10 were sequenced in this study (sequences submitted to GenBank under Accession Numbers JF312218 View Materials – JF312225 View Materials and JF312227 View Materials – JF312228 View Materials ) while P. (P.) lucensis (Issel, 1866) ( AF367651 View Materials , Wilke et al. 2001), P. (P.) macrostoma negropontica (Clessin, 1878) ( EF061915 View Materials , Szarowska et al. 2006), Hydrobia acuta acuta (Draparnaud, 1805) ( AF278804 View Materials , Wilke, Rolán and Davis 2000) and Pyrgula annulata (Linnaeus, 1758) ( AY341258 View Materials , Szarowska et al. 2005) were obtained from GenBank. Hydrobia acuta acuta and Pyrgula annulata were used as outgroups.
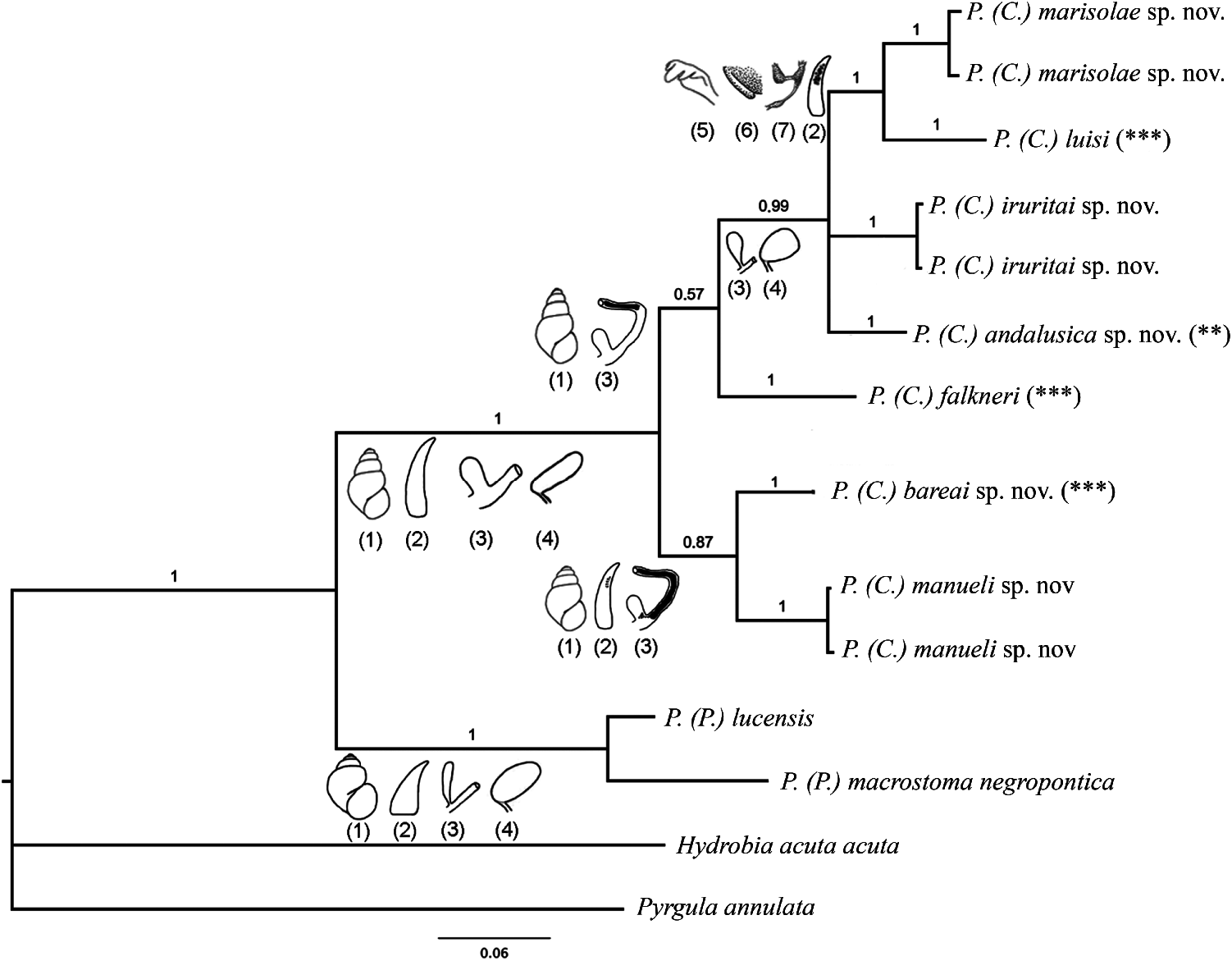
Bayesian phylogenetic reconstruction showed strong support for both the Corrosella subgenus clade and for all the new species here described ( Figure 26 View Figure 26 ). Within the Corrosella subgenus, two clades exist, but with relatively poor posterior probability values: (I) P. (C.) marisolae sp. nov., P. (C.) luisi , P. (C.) iruritai sp. nov., P. (C.) andalusica sp. nov. and P. (C.) falkneri and (II) P. (C.) bareai sp. nov. and P. (C.) manueli sp. nov. Within the first Corrosella clade the relationship between the first four species is well supported but the position of P. (C.) falkneri as a member of this clade has less support.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.