
Pseudamnicola (Corrosella) falkneri ( Boeters, 1970 )
|
publication ID |
https://doi.org/ 10.1080/00222933.2011.623358 |
|
DOI |
https://doi.org/10.5281/zenodo.10536670 |
|
persistent identifier |
https://treatment.plazi.org/id/03C9996E-FFCA-2C22-EB43-89BDFDAAFA63 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudamnicola (Corrosella) falkneri ( Boeters, 1970 ) |
| status |
|
Pseudamnicola (Corrosella) falkneri ( Boeters, 1970) View in CoL
Corrosella falkneri ( Boeters, 1970) View in CoL : p.65–66, figs 1, 3, 5, 8, 10 and 11.
Type locality
According to its original description this species was collected by Falkner on 28 September 1967 in two streams, separated by 120 m, at “Cerro de la Virgen”, Granada , Spain. The eastern flow, between the source and a watering place, is the type locality. The Cerro de la Virgen is located between Galera and Orce above the southern edge of the river Orce ( Boeters 1970). In 1988, Boeters added UTM WG 47 to this type locality .
Type material
Holotype SMF , Paratypes SMF, MP, RNHL, F , BOE 222 et 223 ( Boeters 1970). [ MP (= MNHN), RNHL (= RMNH), F (= FALK or Falkner, according to original writing in collection’ labels)].
Types: Holotype SMF 219026 About SMF , Paratypes SMF 219027 About SMF /1, 219028/10, MNHN, FALK, RMNH, BOE 222 and 223 ( Boeters 1988) .
Material examined
Type material. We examined a photograph of the holotype ( SMF 219026 About SMF ) labelled as “ Abfluss des östlichen Quellaustritts, Cerro de la Virgen, Granada. Falkner leg. 28/9/ 67 ex. BOE 222 ”, five shells from SMF 219028 About SMF / 10 labelled as “ Paratypen, Spanien: Granada: Cerro de la Virgen: W Quellenaustritt, G. Falkner 28.9.67 ( BOE 223 b)”, 10 shells from the NHMW 107332 View Materials labelled as “ Paratypen ex BOE 223 b. Corrosella falkneri Quellenaustritt der W. Balsa, Cerro de la Virgen , Granada FALKNER leg 28 IX 1967 ”, four shells (dry specimens) NHMW 107331 View Materials labelled as “ Paratypen ex BOE 223 b Pseudamnicola (Corrosella) falkneri Westl. Quellenaustritt am Cerro de la Virgen zw. Galera u. Orce oberh. des Süduffers der Vega d. Rio Orce, Granada leg. Falkner 28.9.1967 ”, 15 shells (most of them juveniles) recently collected at 223b (D.M., N.M. and E.M., 20 March 2011, MNCN 15.05 About MNCN /49430) .
All published papers ( Boeters 1970, 1988) mentioned the existence of more than one sample collected at locality 223. The presence of material labelled as “ BOE 223b” in SMF and as “ex BOE 223b” in NHMW alerted us to the existence of a sample 223a. Boeters (personal communication) kindly clarified that the description of P. (C.) falkneri was based on specimens of three samples received from Gerhard Falkner as follows:
- BOE 222 is the type locality (“écoulement oriental”, according to Falkner “Abfluss des östlichen Quellaustritts” [flow from the eastern spring emergence]). Type material from this sample: holotype : SMF 219026 About SMF , paratypes : SMF 219027 About SMF / 1 in addition to BOE collection (Boeters personal communication) that comprises BOE 222 specimens either in alcohol (now BOE 0222 ) or shells (now BOE 2629 ) .
- BOE 223 is the second original locality (“écoulement occidentale”). However G. Falkner distinguished between “Acequia unterhalb der westlichen Balsa” [acequia downhill of the western balsa] ( BOE 223 a) and “Quellaustritt” [spring emergence] ( BOE 223 b). Paratypes at present in BOE collection (Boeters personal communication) comprise animals in alcohol (from BOE 223 a, now BOE 0223 ) and shells (from BOE 223 b, now BOE 2630 ) .
Neither the papers by Boeters (1970, 1988) nor the labels of the collection materials mention the names of the springs, and it was therefore difficult to identify the precise location of the type localities. Boeters (personal communication) provided additional data that have allowed us to locate them. The exact location of the Boeters’ samples is: (European Datum 1950): BOE 222, UTM: 30S 543001 4175819, BOE 223a, UTM: 30S 542838 4175797, BOE 223b, UTM: 30S 542840 4175779 (all explored by D.M., N.M. and E.M. 20 March 2011). Water flows near Cerro de la Virgen are now dry or destroyed and only a few empty shells were found at locality 223b (D. Moreno, personal communication). The area was extensively explored and Pseudamnicola (Corrosella) falkneri was found in five springs at the town of Orce close to Orce river. We accordingly based our anatomical descriptions and molecular data on these specimens. All the material is deposited at the MNCN except for some specimens mentioned below that have been deposited at the NHMW. Localities and collection numbers are as follows: La Armada spring, Orce, Granada, UTM: 30S 0546272/4176139, JM.B., 21 May 2008, MNCN 15.05/49161 (96% ethanol, Figure 8D View Figure 8 and ESEM preparation, Figure 5A,D–G View Figure 5 ,), MNCN /ADN 34847–34851 (96% ethanol), NHMW 92420/ 10 spec. (70% ethanol); JM.B., 30 October 2008, MNCN 15.05/49162 (70% ethanol); Palo spring, Orce, Granada, UTM: 30S 0545712/4175851, JM.B., 21 May 2008, MNCN 15.05/49163 (96% ethanol, Figure 8F View Figure 8 ) and MNCN /ADN 34852–34857 (96% ethanol); JM.B., 30 October 2008, MNCN 15.05/49164 (70% ethanol); Zarza spring, Orce, Granada, UTM: 30S 0544907/4176049, JM.B., 21 May 2008, MNCN 15.05/49165 (96% ethanol) and MNCN /ADN 34858–34859 (96% ethanol); La Pi spring, Orce, Granada, UTM: 30S 0544632/4175976, JM.B., 21 May 2008, MNCN 15.05/49166 (96% ethanol) and MNCN /ADN 34860–34861 (96% ethanol); Las Mimbreras spring, Orce, Granada, UTM: 30S 0545875/4175182, JM.B., MNCN 15.05/49167 (96% ethanol) and MNCN /ADN: 34862–34863 (96% ethanol).
Other material examined. Tubos spring, Castril , Granada, UTM: 30S 0520451/4185742, JM.B., 21 May 2008, MNCN 15.05 About MNCN /49168 (96% ethanol) and MNCN /ADN: 34864–34873 (96% ethanol); spring in Pontezuela , Castril, Granada, UTM: 30S 0519553/4184431, JM.B., 21 May 2008, MNCN 15.05 About MNCN /49169 (96% ethanol) and MNCN /ADN: 34874–34876 (96% ethanol); spring in Castril, Granada, UTM: 30SWH2087, JM.B., 26 August 2006, MNCN 15.05 About MNCN /49170 (96% ethanol) and MNCN /ADN 34877 (96% ethanol); Dos Caños spring, Castril, Granada, UTM: 30S 0520478/4185772, D.D., 13 October 2008, MNCN 15.05 About MNCN /49171 (70% ethanol, Figure 8A View Figure 8 , and ESEM preparation, Figure 5B View Figure 5 ) and MNCN /ADN: 34878–34893 (frozen material); La Errá spring, La Dehesa, Albacete, UTM: 30S 0572400/4242458, D.D., 30 March 2008, MNCN 15.05 About MNCN /49172 (70% ethanol and ESEM preparation, Figure 5C View Figure 5 ) and MNCN / ADN 34894–34898 About MNCN (frozen material); stream in Letur, Albacete, D.M., 12 October 1994, MNCN 15.05 About MNCN /49173 (70% ethanol); Fuente García stream in Cordovilla, Albacete, D.M., 18 May 1997, MNCN 15.05 About MNCN /49174 (70% ethanol); Oria, Almería, UTM: 30S 0570250/4150420, JM. B., December 2009, MNCN 15.05 About MNCN /49198 (96% ethanol) and MNCN / ADN 34965–34966 About MNCN (96% ethanol) .
Material examined for morphometry
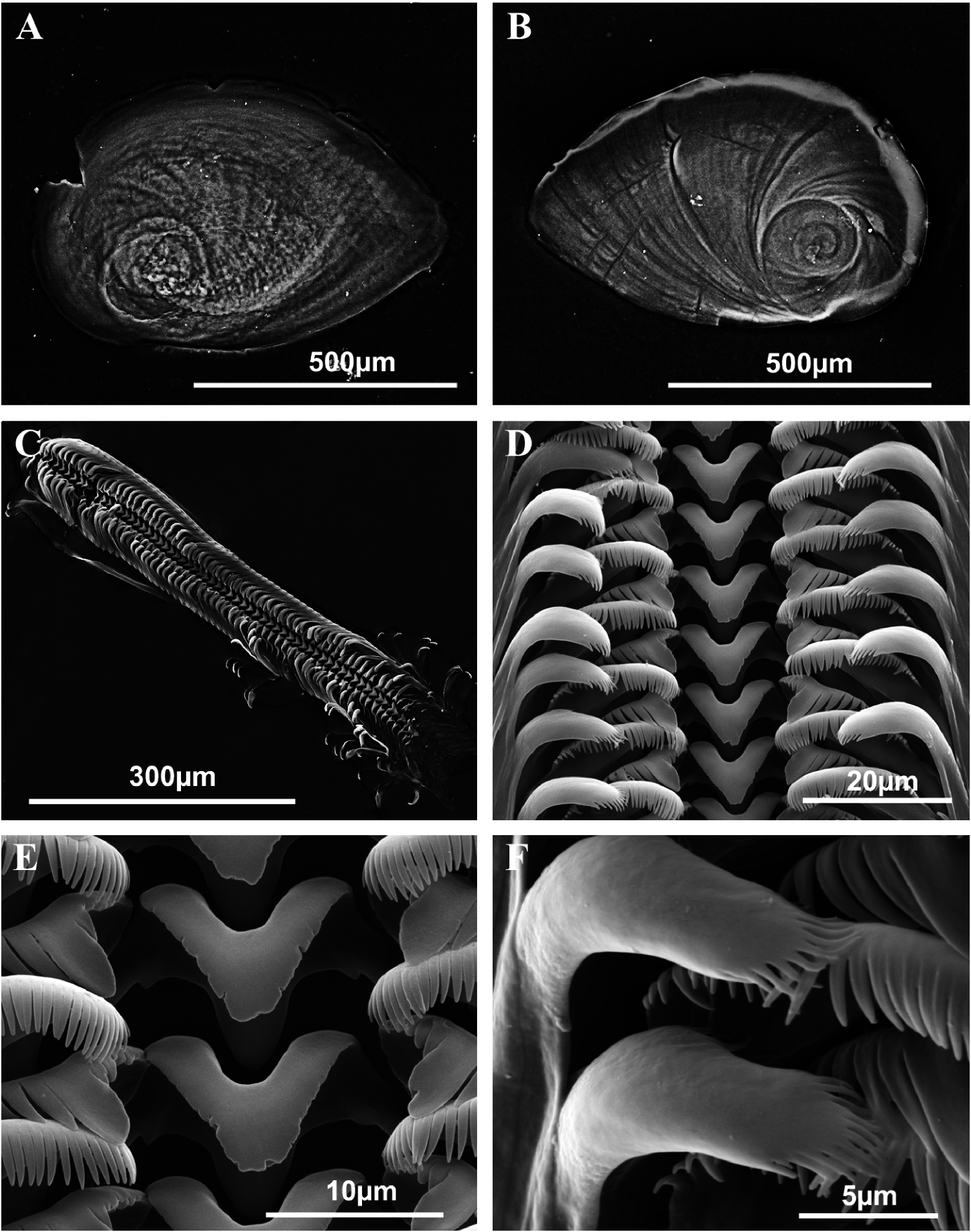
Measurements of the holotype based on published photographs ( Boeters 1970, fig. 10; Boeters 1999, fig. 3) ( Figure 8A View Figure 8 ) and the available paratypes ( Figure 8B,C View Figure 8 ) were compared with measurements of specimens of all the new localities where the species has been found. As explained below, La Armada spring specimens (Orce) were selected for morphometric comparisons with the other species here examined because they were the least eroded and most had a preserved apex. Shell, anatomical, operculum and radular measurements for La Armada (Appendix Tables 1–8) correspond to males and females collected in May and October.
Diagnosis
Shell small with yellowish periostracum; radial sculpture on teleoconch; outer peristome with a slight sinuosity in lateral view; plain central radular tooth weakly serrated and lateral teeth with six lateral cusps at each side of median cusp; bursa copulatrix long cylindrical; seminal receptacle elongated; renal oviduct black pigmented until loop; prostate gland three or four times longer than wide; penis slender tubular with a small black patch of pigment in distal region; nervous system black pigmented with a supraoesophageal connective eight times longer than suboesophageal.
Description
Shell ovate-conic, yellowish periostracum with 4.5–5 spire whorls, height 2–2.6 mm in non-eroded specimens ( Figures 5A–C View Figure 5 , 8D View Figure 8 , Appendix Table 1). In some populations, shells are very eroded at their apex or apex is even lost in different measure showing white colour in this zone; protoconch with around 1.25 whorls, 350 µm total length and nucleus width approximately 100 µm ( Figure 5E,F View Figure 5 ); granular microsculpture with granules more strongly patched at the protoconch nucleus ( Figure 5G View Figure 5 ); teleoconch with radial sculpture marks on its surface, more marked on body whorl which occupies more than half total shell length; peristome orthocline, oval, complete, with a thin outer lip and a thicker inner lip which is in contact with body whorl hiding the umbilicus; outer peristome simple showing slight sinuosity in lateral view ( Figure 5D View Figure 5 ).
Operculum with around 2.5 spire whorls on internal side; oval muscular attachment mark appears near the nucleus ( Figure 6A,B View Figure 6 , Appendix Table 2).
Radula approximately eight times longer than wide ( Figure 6C View Figure 6 , Appendix Table 4) of intermediate size (representing 30% of total shell length); contains around 62 rows of teeth; central tooth flattened, without cusps, border only weakly serrate ( Figure 6D,E View Figure 6 ) with small basal cusp on each side; lateral teeth contain tongue-shaped central cusps and six laterals (external ones longer than internal); inner marginal teeth have 33 cusps which are smaller towards the tooth base, but the fourth cusp from the tooth base is wider and longer than those surrounding it; outer marginal teeth with 25 cusps occupying over 25% of tooth surface ( Figure 6D–F View Figure 6 ).
Pigmentation and anatomy. Head and tentacles dark brown, but pigment absent from ocular lobes ( Figure 7E View Figure 7 ); foot intermediate in size with dark brown pigmentation on its dorsal side. Ctenidium in the middle of the pallial cavity; contains 12–13 gill filaments taller than long; osphradium opposite middle section of ctenidium ( Figure 7C View Figure 7 , Appendix Table 3). Stomach and style sac longer than wide (Appendix Table 5); its anterior region being surrounded by intestine, pigmented in some specimens ( Figure 7F View Figure 7 ); rectum slightly S-shaped in pallial cavity.
Two morphologies of female genitalia observed within the same population: one with both glands in the pallial oviduct approximately equal in size (Appendix Table 6) and a long cylindrical bursa folded into a J-shape ( Figure 7G,H View Figure 7 ), sometimes its distal end also folds; and the other with a capsule gland longer than albumen gland and cylindrical U-shaped bursa copulatrix ( Figure 7I,J View Figure 7 ); in both, the renal oviduct is straight and lacks pigment from its joining point with the bursal duct to oviduct loop, thereafter it becomes blackish and undulates; small elongated seminal receptacle without duct lying on renal oviduct above insertion of the bursa copulatrix duct.
Male genitalia with a three or four times longer than wide prostate gland (Appendix Table 7), and efferent duct entering into the ventral middle-posterior section and deferent duct emerging in the anterior section ( Figure 7D View Figure 7 ); penis intermediate size with base wider than distal region, flattened with a black small patch of pigmentation in distal region ( Figure 7E View Figure 7 ); straight penial duct runs along the external side of penis.
Nervous system black pigmented, with darker pigment on ganglia; cerebral ganglion equal in size approximately joined by 0.11 mm of commissure; supraoesophageal connective is eight times longer than suboesophageal, which usually consists of a simple strangulation; left pleural ganglion usually larger than right ( Figure 7A,B View Figure 7 , Appendix Table 8). Mean RPG ratio 0.49 (moderately concentrated).
Remarks
After examining the available type material, the labels and the original description ( Boeters 1970 and personal communication 2010) one can conclude that Boeters considered that: (1) all the material collected by Falkner between Galera and Orce (three samples BOE 222 , 223 a and 223b) was type material, (2) holotype SMF 219026 About SMF and paratypes SMF 219027 About SMF / 1 (ex. BOE 222 ) plus BOE 0222 and BOE 2629 correspond to the type locality “écoulement oriental”, (3) the other two samples (223a and 223b) are from the second original locality “écoulement occidentale”, (4) specimens SMF 219028 About SMF / 10 and the four specimens NHMW 107331 View Materials are subsamples of the same sample (ex. BOE 223 b) (“Westl. Quellenaustritt”), whereas the provenance of the 10 NHMW 107332 View Materials paratypes is still doubtful because of the contradictory information in their original labels; that is, the location “Quellenaustritt der W. Balsa ” seems to correspond to the third sample 223a, although the writing on the label reads “ex. BOE 223 b” .
Figure 8 View Figure 8 shows the apex erosion that can be observed in both the paratypes and shells of recently collected specimens. Erosion frequently produces loss of first whorls and therefore most length measurements are useless, the remaining variables being too few for a comparative statistical analysis. The population from La Armada is the most similar to the holotype and accordingly it was used for conchological comparisons with the other species in this study (see Statistical analysis).
Populations from Granada exhibit two morphologies of female genitalia within the same population, though genetic studies (unpublished data) indicate no divergence between the two morphotypes (0.00%) that would support two different taxonomic entities (see Discussion). However, all females from Albacete have a large bursa copulatrix and smaller pallial oviduct, the bursa copulatrix being even longer in populations from Albacete than Granada. These morphological inter-population differences are reflected in molecular sequences (0.5% for COI) but are relatively low and can only be taken to indicate inter-population variability. Boeters described an unpigmented penis both in the original description ( Boeters 1970) and in his revision ( Boeters 1988). However, we observed that the distal region of the penis contains a small black patch of pigmentation in all the populations examined.
Molecularly, P. (C.) falkneri represents an independent lineage within the subgenus Corrosella , and its relationship with the other two clades is not well defined ( Figure 26 View Figure 26 ). Morphological similarities with the clade comprising P. (C.) marisolae sp. nov., P. (C.) luisi , P. (C.) iruritai sp. nov. and P. (C.) andalusica sp. nov. are: slen- der shell with same SL/SW ratio (about 1.80, Appendix Table 1), long penis also with the same PL/Head length ratio (about 1.30, Appendix Table 7) and pattern of pigmentation in renal oviduct, starting at the joining point with the bursa duct. Shared characters with P. (C.) manueli sp. nov. and P. (C.) bareai sp. nov. include: small trace of pigment on distal region of penis and shorter seminal receptacle compared with the rest of the studied species. Further, the shape and size of the shell aperture in P. (C.) falkneri are similar to that of P. (C.) hydrobiopsis .
Pseudamnicola (C.) falkneri has numerous autapomorphies because it is the only species of Corosella that shows: a shell as small as 2–2.6 mm, central radular tooth without cusps, six lateral cusps in radular lateral teeth, as few as 12–13 gill filaments, two female genitalia morphologies in some populations, short seminal receptacle (around 0.15 mm), prostate gland as long as almost 2 mm and RPG ratio approaching 0.5 (Appendix Table 8).
In this study, we extend the distribution area both in Granada and in Albacete and the species is cited for the first time from the Almería province. Despite this expansion, P. (C.) falkneri was included as “Vulnerable” in the Red Book of the Invertebrates of Andalusia ( Barea-Azcón et al. 2008).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.