
TESTUDINATA Klein, 1760
|
publication ID |
https://doi.org/ 10.5281/zenodo.5374148 |
|
persistent identifier |
https://treatment.plazi.org/id/A02587A3-4946-EF66-FC92-8274FDA8FA53 |
|
treatment provided by |
Marcus |
|
scientific name |
TESTUDINATA Klein, 1760 |
| status |
|
TESTUDINATA Klein, 1760 (sensu Joyce et al. 2004) Protochelys blakii ( Mackie, 1863) nomen dubium
Chelys (?) blakii Mackie, 1863: 41 , fig. 1.
Testudo stricklandi Phillips, 1871: 182 , diagram 41.
Protochelys stricklandi – Lydekker 1889: 220.
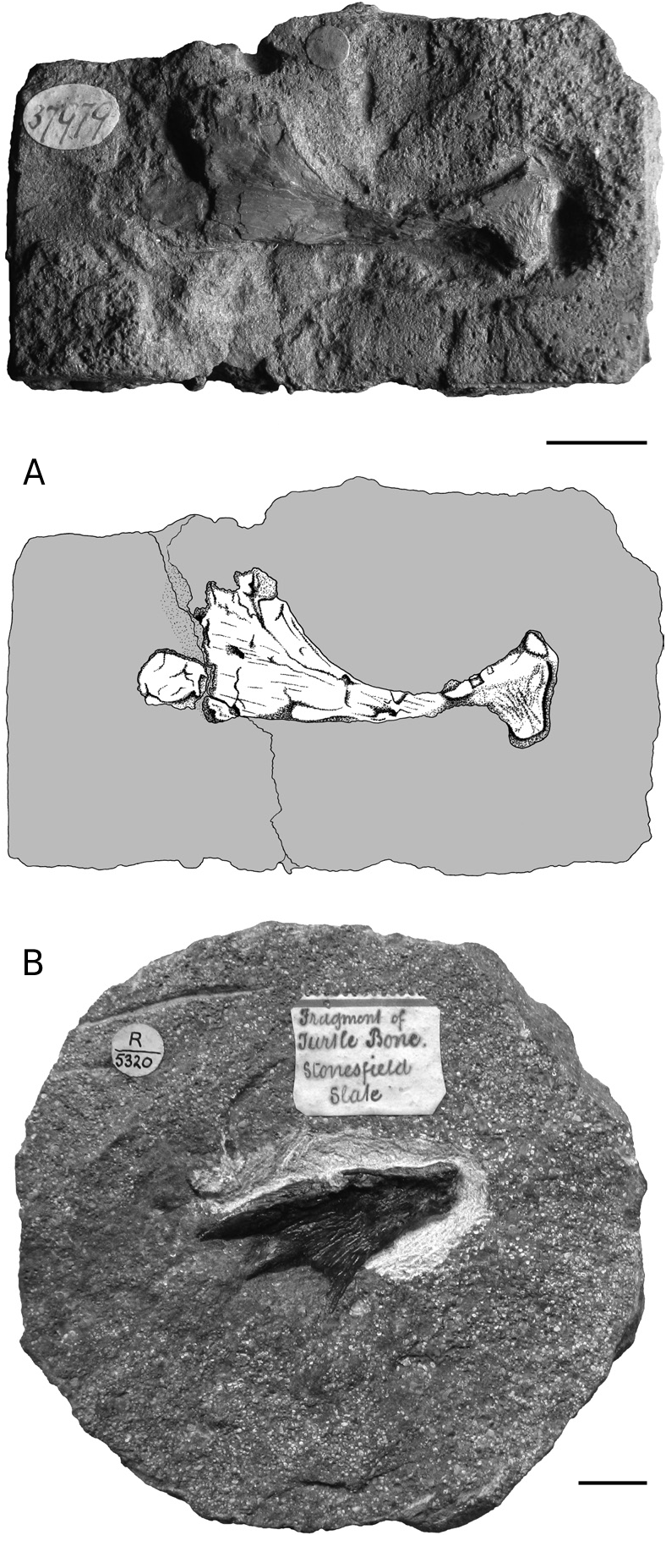
HOLOTYPE. — BMNH 37979 ; an isolated, slightly crushed right coracoid, figured in Mackie (1863) [holotype by monotypy].
REFERRED SPECIMENS. — BMNH 37218, complete vertebral (two slabs); BMNH 37218a, complete vertebral; BMNH R247, half vertebral (two slabs); BM- NH R247a, half vertebral; BMNH 37218b, almost complete vertebral (due to a misreading of the original labelling, the external mould has been erroneously numbered BMNH 39198b after Lydekker [1889]; the correct number is used herein); BMNH R247b, complete pleural (two slabs); BMNH 39198, external mould of a pleural; BMNH 39198a, external mould of a small incomplete scale; BMNH R5320, isolated fragment of plastron; OUMNH J40407 View Materials , complete vertebral; OUMNH J37067 View Materials , complete small vertebral; OUMNH J77375 View Materials + J77376 View Materials , complete vertebral (two slabs, both figured in Phillips [1871]); OUMNH J77377 View Materials , external mould of a small vertebral; OUMNH J77378 View Materials , external mould of a half vertebral.
HORIZON AND AGE. — Stonesfield Slate, Taynton Limestone Formation, Stonesfield, Oxfordshire, England. The Stonesfield Slate is attributed to the lower part of the middle Bathonian ( Torrens 1980; Boneham & Wyatt 1993).
REMARKS
Due to the nature of the specimens, no satisfactory diagnostic characters can be identified. Comparisons with other taxa are also limited (see below). At least as far as the scales are concerned, the consistency of the morphology and preservation suggests that they only represent one taxon. There is no support for the separation of two taxa (one based on the coracoid, the other on the scales) and this would not improve the current taxonomic situation. So, it is sensible to treat all the remains as one species, whose affinities are unknown.
Lydekker (1889: 222) referred to Protochelys the specimen BMNH R896 ( Fig. 1A View FIG ), which he interpreted as a “chelonian scapulo-precoracoid”. In fact, this specimen is not chelonian. It shows pneumatic features and may be interpreted as a possible archosaur cervical rib. The collections of the OUMNH also contain several other specimens misidentified as chelonian:OUMNH J29907 View Materials (a fish scale; Fig. 1B View FIG ) and an unnumbered specimen that consists of a splinter of compact bone (maybe from the base of a crocodilian osteoderm; Fig. 1C View FIG ).
DESCRIPTION
Coracoid
BMNH 37979, as identified by Carter Blake in Mackie (1863), is a right coracoid presented in dorsal view ( Fig. 2A View FIG ). The bone is elongate and slightly crushed. The proximal head is notably enlarged laterally to form the articular glenoid. Medially, the upper part of the sutural surface with the scapula is clearly visible. Posteriorly to the proximal head, the main body of the bone is thin and was probably cylindrical before crushing. Distally, the coracoid expands into a dorsoventrally flattened blade that is characteristic of many turtles. Its posterior margin is broken. The bone may have been concave dorsally but this could be the result of deformation.
Plastron
BMNH R5320 is the only turtle shell bone from the Stonesfield Slate. This is a plastron fragment (hyo- or hypoplastron) probably of the bridge area ( Fig. 2B View FIG ). Three spiny projections are present like those that can be found in embryonic or juvenile individuals or in turtles that do not have a fully ossified shell at adult size. The size of the specimen (40 mm in maximum length) indicates that it does not belong to a hatchling turtle, but it may have belonged to a young individual.
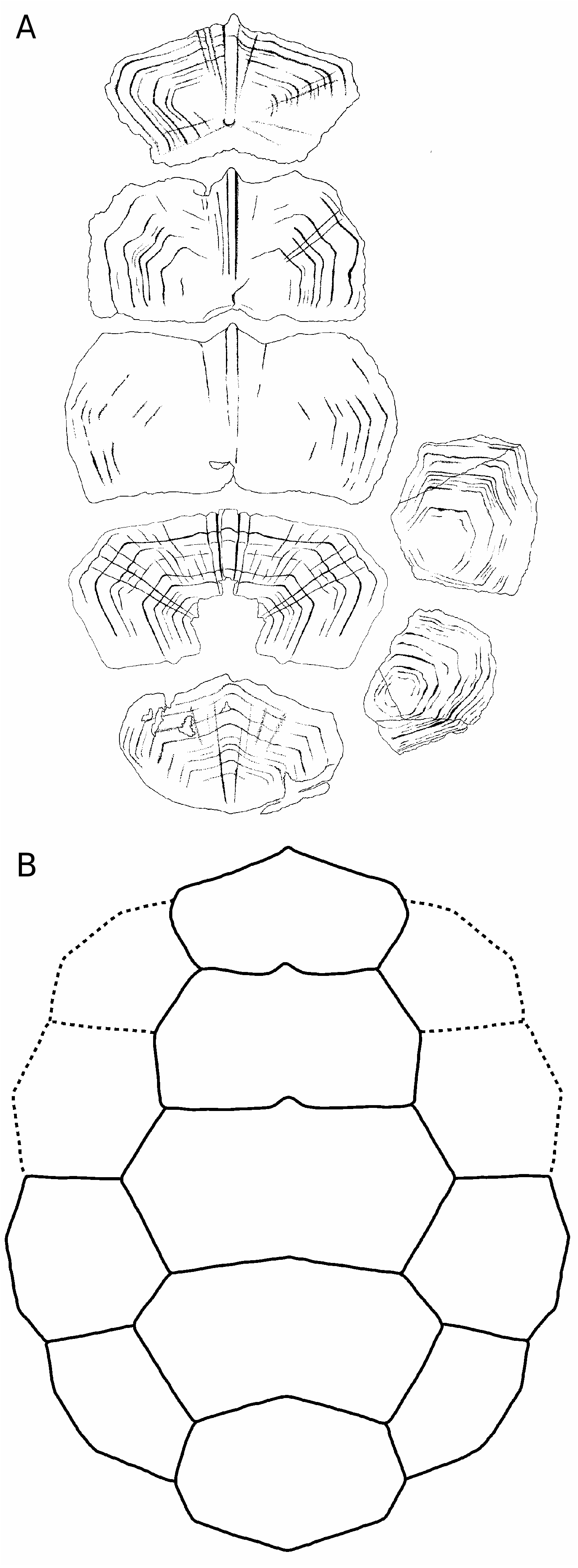
First vertebral scale
BMNH 37218b ( Fig. 3A, B), OUMNH J77375 View Materials + J77376 View Materials ( Fig. 3C, D; see also Phillips 1871: 182, fig. 41.10, 11) and OUMNH J77377 View Materials ( Fig. 3E) are interpreted as first vertebrals because of their symmetrical, pentagonal shape and their concave posterior margin. The bilateral symmetry is underlined by a strong medial keel that is stronger anteriorly and ends abruptly just before reaching the posterior margin of the scale. These scales are almost twice as wide as they are long. It is likely that the second vertebral overlapped the first, as suggested by the presence of a smooth triangular area on the posterior part of the first vertebral and by the disappearance of the medial keel just anterior to this area ( Fig. 3C, D). The anterior margin of the first vertebral is convex and longer than the posterior margin. The long lateral margin faces posterolaterally and contacts the first pleural. On BMNH 37218b, growth rings are poorly preserved in some areas of the scale. A few anteriorly radiating ridges are present on the anteromedial part of the scale. On OUMNH J77375 View Materials + J77376 View Materials , growth rings and radiating ridges are well preserved. OUMNH J77377 View Materials is poorly preserved, but some anteriorly radiating ridges are still visible.
Second vertebral scale
BMNH 37218 ( Fig. 3F, G), OUMNH J77378 View Materials ( Fig. 3H) and OUMNH J37067 View Materials ( Fig. 3I) are interpreted as second vertebrals because of their symmetrical, hexagonal shape and their anterior margin shorter than the posterior margin (generally, vertebrals 2-4 are hexagonal in outline and differ from each other in the relative development of their anterior and posterior margins). These scales have a straight or slightly concave anterior margin and straight posterior margin. They are twice as wide as long (OUMNH J77378 View Materials is incomplete) and have a medial keel. The anterolateral margin contacts the posteromedial margin of the first pleural and faces anterolaterally. The posterolateral margin contacts the anteromedial margin of the second pleural and extends parallel to the anteroposterior axis of the scale. The anterolateral margin is slightly shorter than the posterolateral one. The medial keel protrudes anteriorly from the anterior margin of the scale. The posterior margin presents a medial emargination that probably corresponds to the anterior protrusion of the medial keel of the third vertebral (see below). On BMNH 37218, growth rings are only slightly apparent medially but are still well defined laterally. Some rings are deeply marked but, between those, thinner rings are often present. The spacing between major growth rings is irregular. A few poorly defined anteriorly radiating ridges are apparent laterally. On OUMNH J77378 View Materials , growth rings are better preserved, no intercalated thin rings are present, and the spacing between growth rings is relatively regular. An anteriorly radiating pattern covers the whole surface of the scale. On OUMNH J37067 View Materials , the presence of the medial ridge is uncertain and the majority of growth rings are badly marked (this specimen is poorly preserved). A few anteriorly radiating ridges are present on the anteromedial and posterolateral portions of this specimen.
Third vertebral scale
BMNH 37218a ( Fig. 3J) and BMNH R247a ( Fig. 3K) are interpreted as third vertebrals because of their symmetrical, hexagonal shape and their anterior and posterior margins equal in length. Both specimens exhibit a medial keel that is wider and more pronounced anteriorly. The scale is twice as wide as long with sub-straight anterior and posterior margins. The medial keel protrudes anteriorly from the anterior margin of the scale, whereas a slight medial emargination of the posterior margin probably corresponds to the anterior protrusion of the fourth vertebral. Both lateral margins are oblique with respect to the anteroposterior axis: the anterolateral margin contacts the posteromedial margin of the second pleural, whereas the posterolateral margin contacts the anteromedial margin of the third pleural. The two lateral margins are equal in length. The surface ornamentation of BMNH 37218a is poorly preserved, but growth rings are still well preserved on BMNH R247a. On the latter, thinner (less marked) rings are intercalated between the rather regularly spaced major rings. No radiating pattern is observable, but it should be noted that these specimens are less well preserved than the previously described scales.
Fourth vertebral scale
BMNH R247 ( Fig. 3L, M) is interpreted as a fourth vertebral because of its (original) symmetrical, hexagonal shape and its anterior margin wider than the posterior margin. The right side of the scale and the former emplacement of the embryonic scale are missing. The anterior part of the medial keel is visible on the broken side of the scale. The anterior margin is slightly convex, whereas the posterior margin is slightly concave. Both lateral margins are oblique with respect to the anteroposterior axis: the anterolateral margin contacts the posteromedial margin of the third pleural, whereas the posterolateral margin contacts the anteromedial margin of the fourth pleural. The anterolateral margin is notably shorter than the posterolateral margin. The original scale was approximately twice as wide as long. Due to the preservation, it is unclear whether or not the medial keel protrudes anteriorly, but since a slight medial emargination is present on the posterior margin of the third vertebral (see above) it is probable that it did. Growth rings are relatively well defined and thinner rings are often intercalated between major rings. The spacing between major growth rings is irregular. A pattern of anteriorly radiating ridges is present.
Fifth vertebral scale
OUMNH J40407 View Materials ( Fig. 4A View FIG ) and BMNH 39198a ( Fig. 4B View FIG ) are interpreted as fifth vertebrals because of their symmetrical, pentagonal shape and their convex posterior margin. OUMNH J40407 View Materials has a well-developed medial keel that is wider anteriorly than posteriorly. On BMNH 39198a, an inflexion present on each growth ring near to the broken side of the scale indicates the presence of a keel although this one is not apparent on the specimen (compare with OUMNH J40407 View Materials ). For both specimens, both the anterior and posterior margins are convex, which differs from the condition in other vertebrals. The anterior margin contacts the posterior margin of the fourth vertebral. The long and arched posterior part of the scale in fact presents two separate margins that correspond to the contact with the two last marginals (probably eleventh and twelfth) at the posterior end of the carapace. The short lateral margin faces anterolaterally and corresponds to the contact with the posteromedial margin of the fourth pleural. The scale is wider than long, but proportionally less wide than vertebrals 2-4. Growth rings are well preserved on OUMNH J40407 View Materials , although only slightly defined in some areas. The spacing between major growth rings is irregular and a few anteriorly radiating ridges are present anteromedially. By comparison, only major growth rings are preserved on BMNH 39198a and no trace of a radiating pattern is present.
Pleural scales
BMNH R247b ( Fig. 4C, D View FIG ) is interpreted as a right third pleural because of its non-symmetrical, squarish shape, its straight anterior and posterior margins, its oblique medial margins, and its short posteromedial margin. The scale is slightly longer than wide. The anterior margin is as long as the posterior. On the arched lateral margin, three distinct contacts corresponding to three different marginals can be observed. Growth rings are well preserved, especially on the external mould. Major rings are irregularly spaced and up to three or four less pronounced rings can be seen between some of them. There is no trace of radiating ridges on the scale.
BMNH 39198 ( Fig. 4E View FIG 1 View FIG ) is interpreted as a first or fourth pleural because of its non-symmetrical and irregular pentagonal shape. The original outline of the scale, revealed by growth rings, allows two possible interpretations of this specimen: a right fourth pleural ( Fig. 4E View FIG 2 View FIG ) or a left first pleural ( Fig. 4E View FIG 3). The following observations weaken the second hypothesis: first pleurals are usually more elongate; growth rate should have been greater anteriorly; and the posterior margin that contacts the second pleural should be sub-perpendicular to the anteroposterior axis of the scale. Consequently, we favour an interpretation of BMNH 39198 as a right fourth pleural. The scale is longer than wide. The anteromedial margin (that contacts the posterolateral margin of the fourth vertebral) is damaged and consequently the anterior margin (that contacts the posterior margin of the third pleural) appears shorter than it was originally. The short posteromedial margin corresponds to the contact with the lateral margin of the fifth vertebral. The lateral margin is divided into three parts corresponding to the contact with three different marginals. Growth rings are relatively well preserved. Major rings are irregularly spaced. Few less pronounced rings are present but difficult to distinguish. There is no radiating pattern on the scale.
COMPARISON
It is difficult to compare the turtle scales from the Stonesfield Slate with other taxa because no directly comparable material (i.e. fossil epidermal scales) is known. When describing bony shells, systematists can only access information on the general outlines of scales and mainly use their relative sizes for diagnostic purposes. At Stonesfield, although they match each other fairly well ( Fig. 5A View FIG ), the scales are isolated and obviously pertain to different individuals: they show different growth rates, different sizes and there is no indication that all of the scales come from the same stratigraphical horizon. So that the relative sizes of the Stonesfield scales cannot be assessed. However, the detailed description provided herein enables some comparisons to be made.
A tentative reconstruction of the carapace of the Stonesfield turtle is provided in Figure 5B View FIG . An important characteristic of this turtle is that vertebrals are wider than long, whereas pleurals are slightly longer than wide. This is a rather primitive scheme commonly found in stem turtles and basal crowngroup turtles. More derived turtles tend to have vertebrals that are longer than wide and pleurals that are wider than long.
Among turtles with wide vertebrals and narrow pleurals, the Stonesfield turtle more closely resembles the stem turtles Kayentachelys aprix Gaffney, Hutchison, Jenkins & Meeker, 1987 ( Gaffney et al. 1987), Indochelys spatulata Datta, Manna, Ghosh & Das, 2000 ( Datta et al. 2000), Heckerochelys romani Sukhanov, 2006 ( Sukhanov 2006) and Kallokibotion bajazidi Nopcsa, 1923 ( Gaffney & Meylan 1992), the panpleurodire Notoemys laticentralis Cattoi & Freiberg, 1961 ( Fernandez & Fuente 1994; Lapparent de Broin et al. 2007), and the pleurosternid Dinochelys whitei Gaffney, 1979 ( Gaffney 1979; Brinkman et al. 2000). The aforementioned species share the following features with the Stonesfield turtle: large vertebral 1 with a convex anterior margin that is at least as wide as the posterior margin; vertebral 2 with a posterolateral margin more or less parallel to the anteroposterior axis of the shell; vertebral 4 with a significantly short anterolateral margin; vertebral 5 wider than long with an anterior margin as wide as the posterior margin. The Stonesfield turtle differs from each of these species by several of its characters. Vertebrals 2-4 of Kayentachelys and Indochelys are wider than those of the Stonesfield turtle, whereas they are narrower in Kallokibotion. In Kayentachelys , vertebral 1 is at least three times wider than long and has three distinct margins anteriorly (one for each marginal 1 and one for the cervical scale). Vertebral 1 of Dinochelys and Notoemys is also wider than that of the Stonesfield turtle. In Kallokibotion, vertebral 1 is narrower and more elongated than that of the Stonesfield turtle and its lateral margin is parallel to the anteroposterior axis of the shell. In Notoemys , the lateral margin of vertebral 1 are also parallel to the anteroposterior axis of the shell and all intervertebral sulci are straight. Dinochelys also has straight intervertebral sulci. Heckerochelys differs from the Stonesfield turtle in having a longer vertebral 2, vertebral 3 with strongly concave posterior margin and vertebral 4 with only one lateral margin.
Concerning the scale ornamentation, none of the aforementioned species is known to have one, although it is possible that scale ornamentation does not always leave a trace on the bone plates of the shell. One can notice that the anterior margin of vertebrals 2-5 of Kayentachelys presents a welldefined, short medial protrusion similar to that of the Stonesfield turtle. Other turtles known to have a scale ornamentation of anteriorly radiating ridges on the vertebrals (e.g., Proganochelys , Platychelys , Euryaspis , Desmemys , some plesiochelyids and chengyuchelyids) do not match the morphology of the Stonesfield turtle.
Romer (1956, 1966) and Bergounioux (1955) referred the turtle from Stonesfield to the Pleurosternidae . Pleurosternon bullockii ( Owen, 1842) , Glyptops plicatulus (Cope, 1877) and “ Glyptops ” typocardium (Seeley, 1869) ( Gaffney 1979; Milner 2004) all have vertebrals that are reduced in width and pleurals wider than long. Among pleurosternids, only the Late Jurassic Dinochelys whitei has a scale pattern somewhat similar to that of the Stonesfield turtle. However, as it is also true for other species ranging from Early Jurassic stem turtles to basal panpleurodires (see above), one can only assume that this scale pattern is plesiomorphic. In other words, a referral to the pleurosternids is not better supported than a referral to the stem Testudines or the Panpleurodira in the current state of knowledge.
TAPHONOMY
The preservation of turtle epidermal scales is rather uncommon in the fossil record. We are aware of only two other cases: a shell fragment from the Pleistocene of NE Thailand (J. Claude pers. obs.) and a shell of Neurankylus from the early Paleocene of New Mexico ( Sullivan et al. 1988). In the later case, the scales are preserved on approximately 85% of the carapace, are rich in iron and manganese, and are apparently preserved with the original colour pattern ( Sullivan et al. 1988: fig. 3). In both cases, the scales are preserved as a thin layer on the surface of the bony shell, but the Stonesfield specimens are the only known example of isolated scales in the fossil record. Isolated epidermal scales can result from two processes: shedding during growth or post mortem disarticulation.
To our knowledge, no detailed studies of the post-hatching development of shell scales have been published. In a recent review on the use of growth rings to estimate turtle age, Wilson et al. (2003) discuss this lack of literature on epidermal scale growth.Two patterns are seen in extant turtles ( Wilson et al. 2003: fig. 1). In some turtles (e.g., Terrapene carolina (Linnaeus, 1758)) , scales do not shed and old layers are retained on the external surface of new ones. In such cases, growth rings correspond to the superposition of successively larger and younger scales. Such scales become thicker with age, although old layers are often progressively worn by abrasion. In other turtles (e.g., Trachemys , Orlitia , Chrysemys ), the old scale is shed while the new one grows beneath it. Although the mechanism is not yet understood, the old layers leave an impression, corresponding to the growth ring, on the new scale. Shed keratinous scale layers are thin, translucent and flexible, and are consequently unlikely to fossilise. Moreover, we were unable to find any reference mentioning the persistence of shed scale layers in the environment, which might suggest that they are quickly destroyed after shedding.
The isolated nature of Stonesfield scales probably results from disarticulation rather than shedding: shed scales are thinner and more fragile than complete scales, and are therefore less likely to fossilise. Moreover, shed scales would probably not display such marked growth rings. Indeed, extant shedding turtles tend to have smooth epidermal scales with poorly developed rings.
Turtle taphonomy is poorly studied and the few available studies deal primarily with bones, which is little help in case of the Stonesfield material. Frustratingly, studies on extant species often fail to mention epidermal scales at all (see Brand et al. 2003 for a more detailed review). Howev- er, among these neontological studies, Bourn & Coe (1979) reported the disarticulation sequence of Geochelone gigantea (Schweigger, 1812) (the Aldabra tortoise) and stated that, at some point in the sequence, scales detach themselves from each other and from the bony layer before falling. Dodd (1995) documented the disarticulation patterns of 80 turtle carcasses, representing three families (emydids, testudinids and trionychids), in a terrestrial sandhill habitat in North Florida. As noted by Bourn & Coe (1979), Dodd (1995) also observed that the keratinous scales of emydids and testudinids eventually detached themselves individually from the shell (trionychids have no scales but a leathery skin covering the shell). He noted that the vertebrals and pleurals generally detach simultaneously, or the former closely followed by the latter. Marginals are often the first group to detach, although they persist longer than the vertebrals and pleurals in the testudinid Gopherus polyphemus (Daudin, 1802) . Dodd (1995: 383) observed that disarticulated scales may remain close to the carcass for an extended period. Brand et al. (2003) carried out an experimental study to compare the disarticulation processes for the common slider turtle ( Trachemys scripta (Schoepff, 1792)) in different environments. They observed that separation between scales and shell bones occurred at the same time as shell disarticulation.Unfortunately, the detached elements were removed immediately from the experimental area so that there was no estimation of the time that disarticulated scales could remain in the environment.
Thus, taphonomic studies on turtles have not so far explained the fate of scales once detached or shed. For example, it is unknown how long keratinous scales can remain in the environment, how they are affected by transport or what conditions may be conducive to their preservation. The Stonesfield specimens are the only isolated fossil turtle scales known to date, though it is possible that others exist and have been misinterpreted or remain unrecognised and undescribed. The preservation of isolated keratinous elements is unusual in the fossil record and probably requires particular burial environments and transport conditions. In addition, there are currently no detailed taphonomical studies of the Stonesfield biota in general that could explained the preservation of these scales, although similarities of preservation with the plant remains from the same locality are striking ( Cleal & Rees 2003).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
TESTUDINATA Klein, 1760
| Anquetin, Jérémy & Claude, Julien 2008 |
Protochelys stricklandi
| LYDEKKER R. 1889: 220 |
Testudo stricklandi
| PHILLIPS J. 1871: 182 |
Chelys
| MACKIE S. J. 1863: 41 |