
Postaphanostoma nilssoni, Kånneby, Tobias & Jondelius, Ulf, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3736.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:0D412548-2436-434E-93F7-7D2B4EAB53CB |
|
DOI |
https://doi.org/10.5281/zenodo.5658312 |
|
persistent identifier |
https://treatment.plazi.org/id/EB2DA62F-5649-FFB0-37AF-25EAABCBE18D |
|
treatment provided by |
Plazi |
|
scientific name |
Postaphanostoma nilssoni |
| status |
sp. nov. |
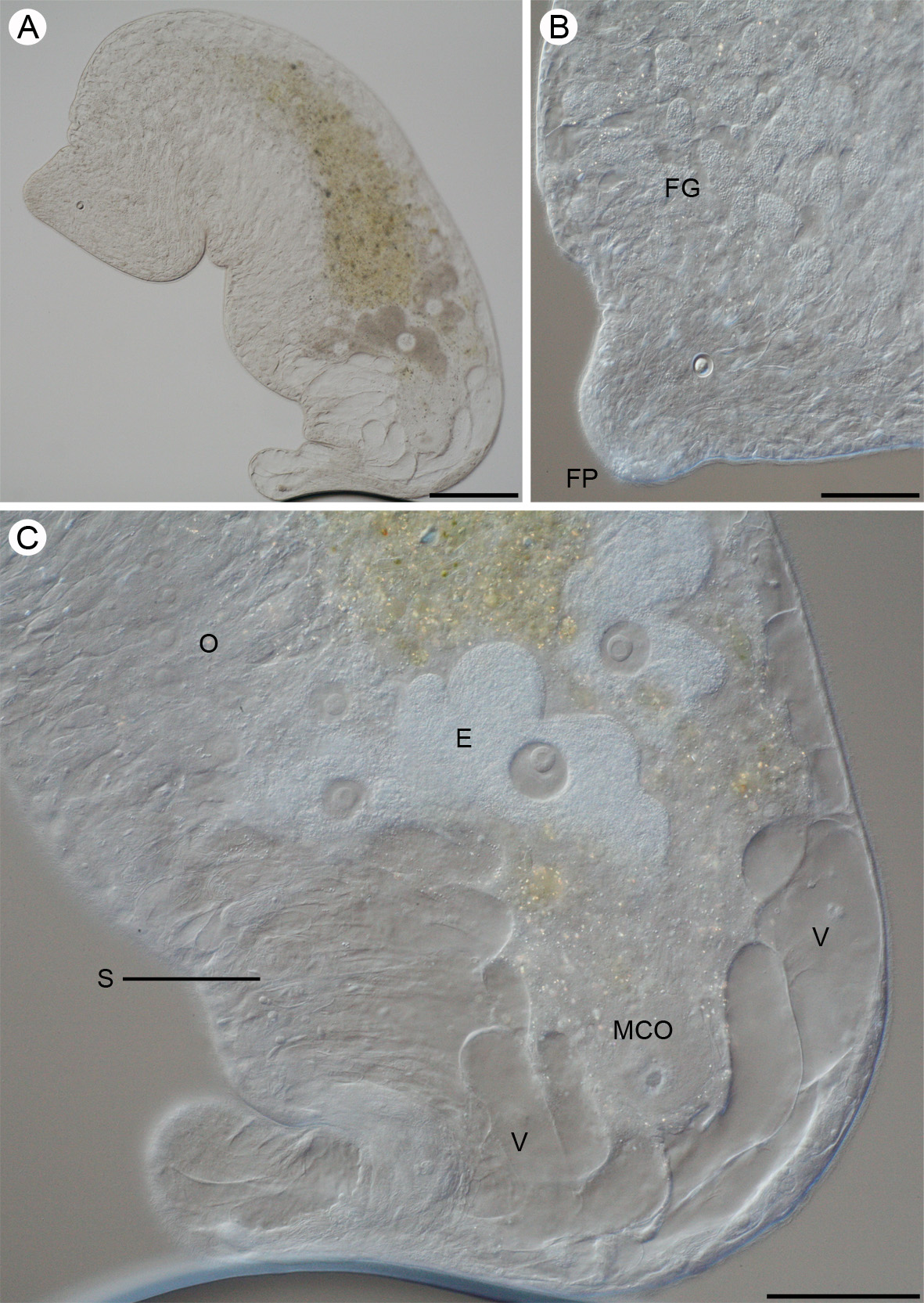
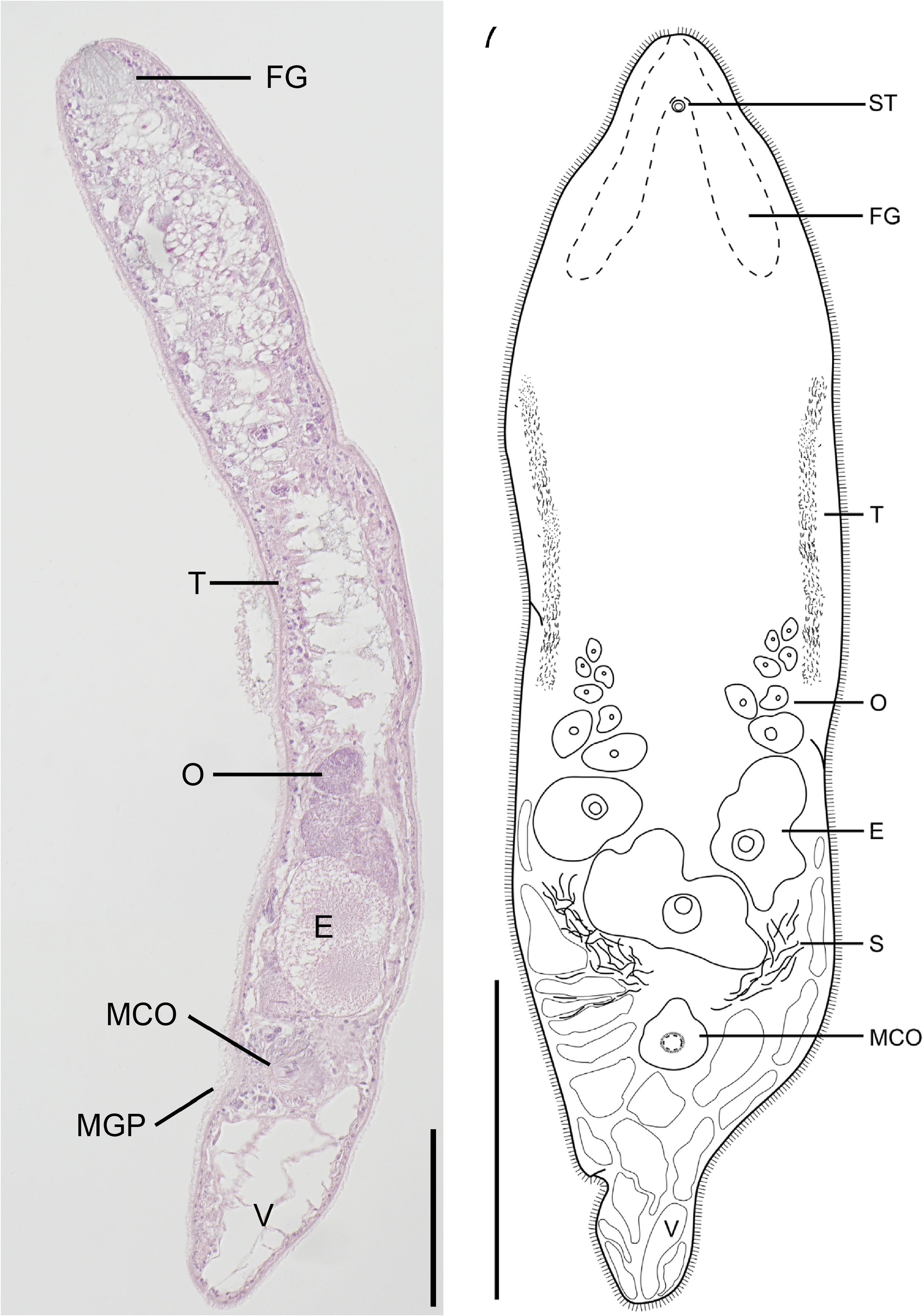
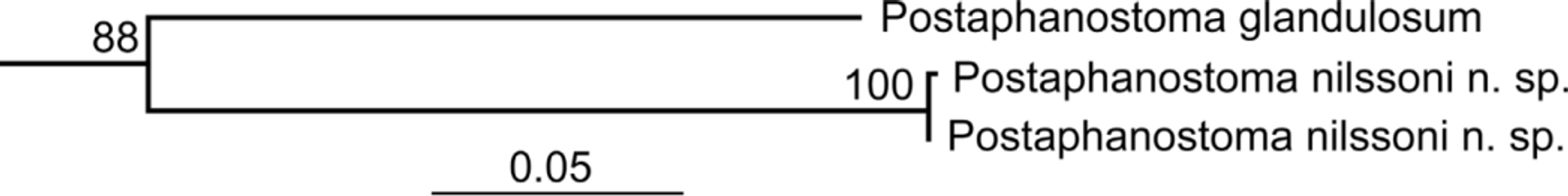
Postaphanostoma nilssoni n. sp. ( Figs. 5–8 View FIGURE 5 View FIGURES 6 – 7. 6 View FIGURE 8 )
Type Material: Holotype—SMNH Type-8464 (sagittal sections); Paratype—SMNH Type-8465 (longitudinal sections); Paratype—SMNH Type-8466 (sagittal sections).
Type Locality: Rock pool by Estación Costera de Investigaciones Marinas (ECIM), Las Cruces, Chile (33° 30' 10.548" S, 71° 37' 54.012" W), intertidal in shell gravel.
Other Material Examined: Whole mounts of living specimens.
Etymology: This species is named in honor of Karin Sara Nilsson who helped collecting the material.
Description: Body cylindrical, variable in size and shape. Living free-swimming specimens 4–5 mm in length. Living mounted specimens 2–3 mm and fixed specimens approximately 1 mm in length. Width varies between 0.6–1.25 mm in living highly compressed mounted specimens and is approximately 100 µm in fixed specimens. Anterior and posterior ends rounded with the posterior end usually narrower ( Fig. 5 View FIGURE 5 A). Body colorless, unpigmented. Central syncytium is yellowish and eggs are of a brownish color ( Fig. 5 View FIGURE 5 A). Rhabdoid glands small (appr. 10 µm long), scattered in the anterior end of the body.
Epidermis completely ciliated, approximately 3–4 µm in sagittal sections ( Fig. 6 View FIGURES 6 – 7. 6 ). Body wall apparently with well developed dorsoventral musculature. Large vacuoles, up to 300 µm in diameter, confined to posterior part of body ( Figs. 5 View FIGURE 5 A, C). Smaller vacuoles are also present along the lateral sides of the posterior third of body ( Fig. 7 View FIGURES 6 – 7. 6 ).
Statocyst 25 µm in diameter and located 160 µm from the anterior end at U 6 in mounted specimens. Statolith 13 µm in diameter. Frontal organ well developed with numerous frontal glands, extending from anterior end along the lateral sides to U18 ( Figs. 5 View FIGURE 5 B; 7).
The position of the mouth could not be determined. Central syncytium, yellowish, extends from U23 to the mature eggs at U64. From U23 to U40 the central syncytium is diffuse but then becomes more compact ( Fig. 5 View FIGURE 5 A).
Ovaries paired situated anterior to male copulatory organs. They extend from the middle of the body at U50 to the anterior parts of the male copulatory organ at U76 ( Figs. 5 View FIGURE 5 A, C; 6). Bursa and female gonopore absent. Testes paired, extending along the lateral sides of the body, from U26 to U51 ( Fig. 7 View FIGURES 6 – 7. 6 ). Seminal vesicle absent. Sperm is collected in false seminal vesicles, a collection of sperm not surrounded by a muscular wall (see Westblad 1945; Faubel 1976; Ehlers & Dörjes 1979). The male copulatory organ is formed by an invagination of the ventral epidermis ( Fig. 6 View FIGURES 6 – 7. 6 ). The wall of the male copulatory organ is muscular and surrounded by eosinophilic gland cells ( Fig. 6 View FIGURES 6 – 7. 6 ). Male gonopore located at U78, between the large posterior vacuoles and the mature eggs.
Diagnosis and taxonomic remarks: Postaphanostoma nilssoni n. sp. can be separated from all other species within the genus based on its large body size. The following morphological characters identifies the new species as a member of Postaphanostoma : (i) a well developed frontal organ with numerous frontal glands; (ii) small poorly developed rhabdoid glands; (iii) seminal vesicle absent; (iv) the male copulatory organ is an invaginated continuation of the epidermis and (v) sperm are collected in false seminal vesicles (Dörjes 1968). The maximum likelihood analysis of the concatenated 18S, 28S and COI sequences shows a sister group relationship between P.
nilssoni and P. glandulosum Dörjes, 1968 ( Fig. 8 View FIGURE 8 ). However, P. glandulosum is the only other species within the genus for which we currently have sequences available.
Postaphanostoma atriomagnum Dörjes, 1968 , Postaphanostoma filum Dörjes 1968 and Postaphanostoma glandulosum are all less than 1.5 mm in total body length and only 0.1–0.2 mm in total body width compared to 4– 5 mm and 0.6–1.25 mm in total body length and width respectively, for living specimens of P. nilssoni n. sp. The other species within the genus all possess a seminal bursa and unpaired ovaries while a seminal bursa is absent and the ovaries paired in P. nilssoni n. sp. Moreover the posterior end of the new species contains more vacuoles than in any of the other species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |