
Pirabasoporella baitoae, Zágoršek, Kamil, Ramalho, Laís V., Berning, Björn & Távora, Vladimir De Araújo, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3838.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:458183F5-78CF-4E00-AEA3-8C4477CF829B |
|
DOI |
https://doi.org/10.5281/zenodo.5617692 |
|
persistent identifier |
https://treatment.plazi.org/id/E46D87BE-FF90-FFD3-50A9-FB05FF04FA36 |
|
treatment provided by |
Plazi |
|
scientific name |
Pirabasoporella baitoae |
| status |
sp. nov. |
Pirabasoporella baitoae View in CoL n. sp.
( Figure 5 View FIGURE 5 )
Etymology. Alluding to the Early Miocene type Baitoa Formation in the Dominican Republic.
Material examined. Holotype: USNM 595120, the colony fragment depicted in Fig. 5 View FIGURE 5 A, B, from the type locality at Rio Yaque del Norte; type stratum Baitoa Formation, Early Miocene, (sensu Saunders et al. 1986), Dominican Republic. Paratypes: USNM 595121 to 595125, 5 separate colony fragments. Additional material: More than 100 colony fragments in USNM, numbers NMB 17285–17289 and NMB 16935, NMB 17184 and Olsson 179.
Diagnosis. Pirabasoporella with relatively few frontal-shield pores that become progressively closed during ontogeny, surface often nodular. Apertural pseudosinus relatively wide. Abfrontal rhizoidal pores large and encircled by an elevated rim, strut pores very large. Adventitious avicularia present on abfrontal side of some zooids; apart from the suboral avicularium additional frontal avicularia are lacking. Ovicells unknown.
Description. Colony rigid, reticulate, with bifurcating branches connected by kenozooidal struts, branches growing more or less parallel to each other ( Fig. 5 View FIGURE 5 A, C); struts irregularly developed, jointly budded laterally from strut pore-plate, rarely one strut budded from two adjacent plates ( Fig. 5 View FIGURE 5 G), slightly curved or straight, surface smooth or with faint ridges and a row of slit-like areolar pores near each branch contact. No fenestrules preserved.
Autozooids elongate-pentagonal, widest at about mid-distance, arranged in two alternating longitudinal series, separated by thin raised ridges that are especially prominent on the abfrontal side, the lepralioid part of the frontal shield perforated by up to 40 pores, many of which are closed during ontogeny while the lateral areolar pores remain open, surface occasionally nodular; umbonuloid part confined to distalmost frontal shield. Interzooidal communication via multiporous pore plates. Primary orifice immersed and ill-defined, with inconspicuous condyles, secondary orifice relatively large ( Fig. 5 View FIGURE 5 B), suborbicular, about as wide as long, with a broadly U-shaped pseudosinus comprising 1/5 of total orifice length, formed by two extremely short pointed lateral denticles.
Two types of avicularia present: each autozooid on frontal side with a single, small, suboral avicularium situated directly proximolateral to pseudosinus ( Fig. 5 View FIGURE 5 B), oblique to frontal plane, rising distally, rostrum semielliptical, directed laterally towards median line, cystid raised distally to produce a suboral umbo, crossbar complete, without columella. Some zooids have an additional, slightly larger (see Table 2 View TABLE 2 for measurements) adventitious avicularium on abfrontal side, situated proximal to rhizoidal pore on a somewhat elevated cystid, elongate- oval with complete crossbar ( Fig. 5 View FIGURE 5 F, G)
Abfrontal side of colony relatively smooth to distinctly nodular, zooidal borders marked by prominent ridges with a deep central suture, each zooid with an extremely large oval to circular rhizoid pore (up to 268 µm diameter) positioned relatively distal, surrounded by a thick prominent rim and sealed at its base by a multiporous pore plate with the large pores situated along the margin ( Fig. 5 View FIGURE 5 F). Rhizoid pores of a similar type, but prominently oval ( Fig 5 View FIGURE 5 G), present at distal edge of lateral walls of zooids that do not produce a kenozooidal strut, the pores in these usually smaller, not limited to pore-plate margin.
Ovicells and ancestrula not observed.
Remarks. Pirabasoporella baitoae n. sp. differs from P. atalaiaensis n. sp. and P. chipolae n. sp. in having generally fewer pores in the frontal shield (which occasionally displays a nodular surface), a broader apertural pseudosinus and the occasional presence of abfrontal avicularia while large frontal avicularia are absent. Further differences from P. atalaiaensis are the shorter but broader autozooids, and distinctly larger abfrontal rhizoidal pore plates that are circled by an elevated rim. In contrast to P. c hi po l ae, P. baitoae does not have oral spines.
Among the approximately 100 colony fragments studied, none contained ovicells or indications of their presence. Insofar as P. atalaiaensis , like all other jaculinid species, produces hyperstomial ovicells, we do not interpret their lack in P. ba i t o ae as an indication of internal brooding but perhaps reflecting a response or adaptation to environmental conditions, in which asexual colony growth may prevail over sexual reproduction. A similar strategy is found in Metrarabdotos , for example, in which several hundred autozooids may be formed for every gonozooid ( Cheetham et al. 2001). However, we have no evidence (e.g. repair of broken zooids) that propagation in P. baitoae took place predominantly via fragmentation of colonies. On the other hand, neither ancestrulae nor early astogenetic colonies have ever been reported in any living or fossil jaculinid.
In one instance ( Fig. 5 View FIGURE 5 E) a connecting strut was observed that originated from two different pore plates (i.e. zooids) on one side of the branch while fusing towards, and connecting to, a single pore plate on the opposite branch.
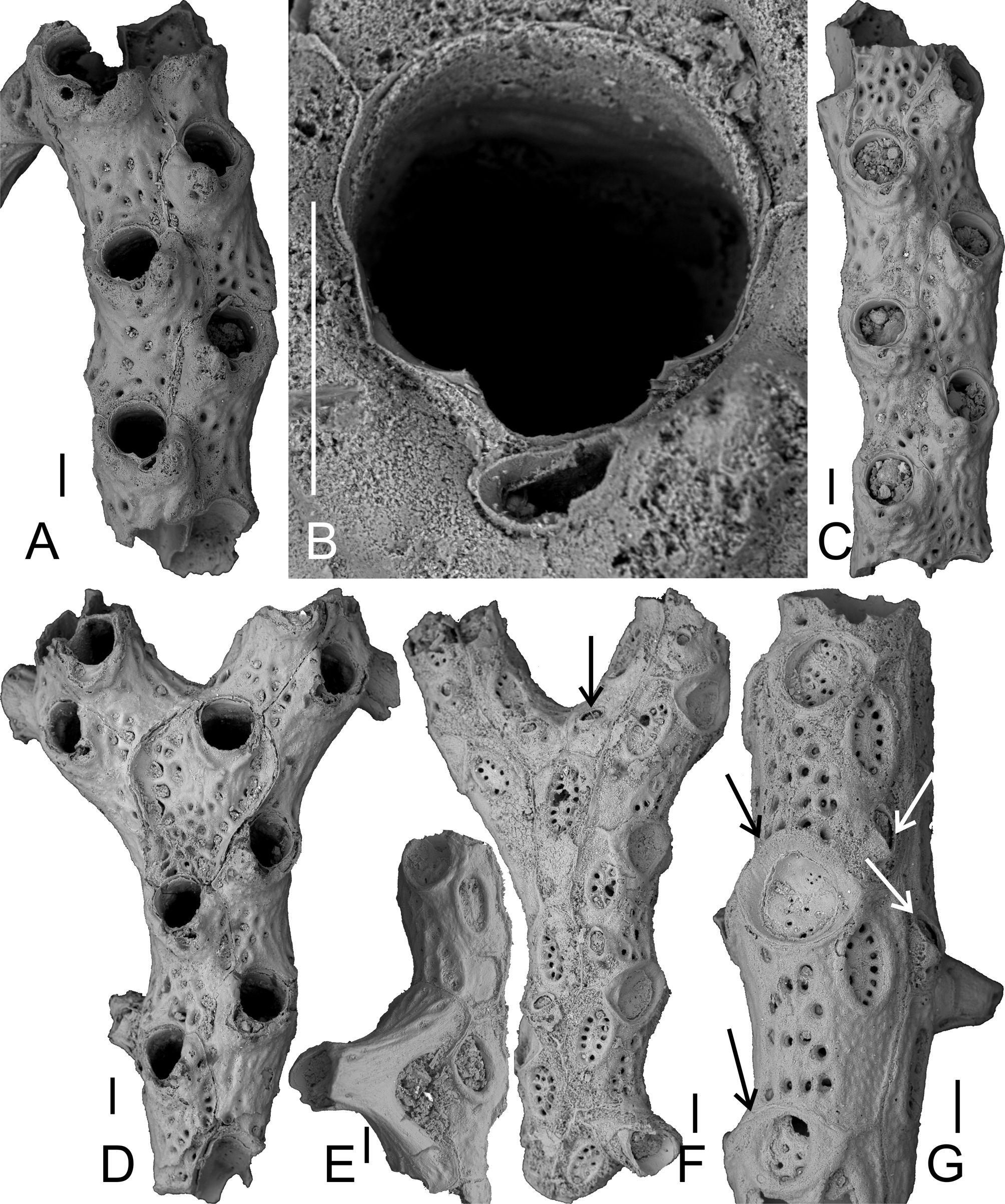
TABLE 2. Measurements for Pirabasoporella baitoae n. sp.
| BW | SD | AL | AW | AD | SPL | SDW | PRL | RPW | AAL | SAL | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| mean | 458 | 183 | 500 | 236 | 151 | 245 | 233 | 186 | 97 | 93 | 57 |
| min | 403 | 153 | 443 | 200 | 142 | 181 | 179 | 128 | 85 | 64 | 44 |
| max | 514 | 202 | 543 | 28 | 158 | 302 | 268 | 219 | 113 | 113 | 81 |
| # | 37 | 18 | 32 | 32 | 64 | 12 | 12 | 24 | 24 | 6 | 5 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |