
Pichidamas piru, Bulnes & Grau & Carbayo, 2018
|
publication ID |
https://doi.org/ 10.1080/00222933.2018.1538468 |
|
publication LSID |
lsid:zoobank.org:pub:04423258-DB01-4CE3-A474-EF6B563C605C |
|
persistent identifier |
https://treatment.plazi.org/id/03C987C0-FFA4-FFA6-2BF3-56CFFC08FEA2 |
|
treatment provided by |
Carolina |
|
scientific name |
Pichidamas piru |
| status |
sp. nov. |
Pichidamas piru View in CoL sp. nov.
( Figures 1–6 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 )
Material examined
Holotype. ZMB 11682. Pichidamas, Entre Lagos municipality, Osorno province , Chile, 40°43'S 72°49'W J. Pérez-Schultheiss Coll., 5 October 1999. Cephalic region: transverse GoogleMaps
sections on 16 slides; pre-prepharyngeal region: frontal sections on three slides; prepharyngeal region: transversal sections on five slides; copulatory apparatus: sagittal sections on 14 slides.
Type locality
Pichidamas, Osorno province, Entre Lagos municipality, Chile; inside a fallen tree trunk. The locality is covered by inner temperate laurifoliate forest dominated by Nothofagus dombeyi and Eucryphia trees . The specimen was collected in a disturbed area, used extensively in livestock foraging, where the original forest alternates with small pastureland patches composed mainly by Holcus lanatus and Agrostis tenuis .
Etymology
The specific epithet ‘piru’ (gender: masc) refers to the word in Mapudungun language, used by the Mapuche (originary people of central Chile) for worm.
External morphology
Body of medium size, after fixation 21 mm long, 2.5 mm wide, 1.4 mm thick ( Figure 1 View Figure 1 (a)). Parallel body margins throughout most of the body length and tapering abruptly towards both anterior and posterior end, both endings rounded. Dorsum background colour light brown, with randomly arranged elongated dark brown patches, giving the appearance of a tree rim ( Figure 1 View Figure 1 (b, c)). Over this pattern, a distinct middorsal dark brown band (0.30 mm) and a faint submarginal stripe (0.28 mm) on each side of the body, external to it there is a rind devoid of pigment. The submarginal stripes extend from the anterior end, up to the posterior end, while the median band commences at 2.4 mm from the anterior tip, extending to the posterior end. The submarginal stripes narrow in their posterior 1/4 of body length, while the middle stripe retains its full width. Ventrally the body is whitish.
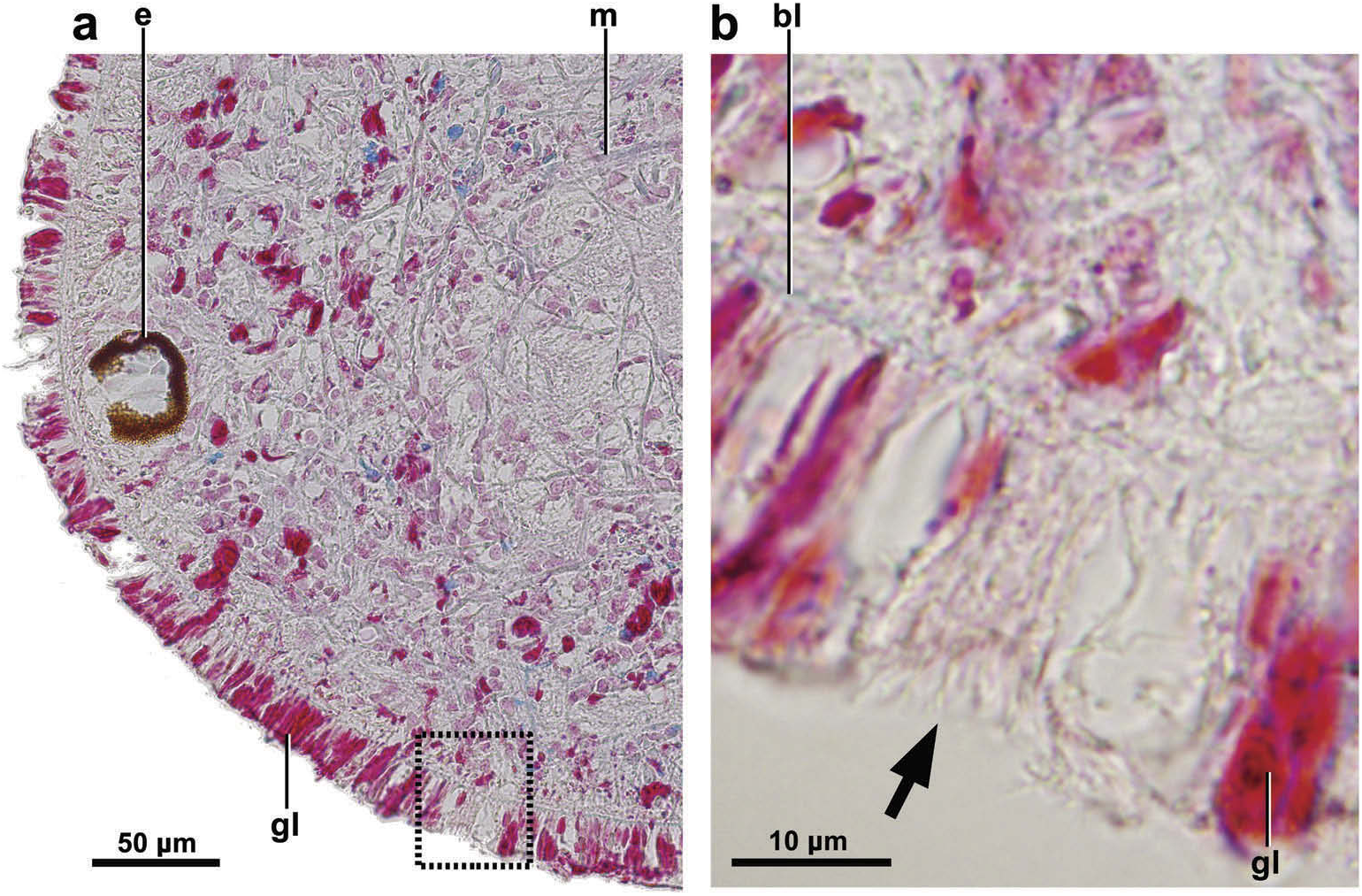
Eyes present from the anterior tip to the posterior end in a single irregular marginal row. At the anterior tip and anterior body margin the eyes are most densely packed ( Figure 1 View Figure 1 (a, b)). The size of the eyes varies between 30 and 80 µm. Sensory pits absent. Instead, spots of cilia in slight epithelial depressions located in the ventrolateral region of the anterior portion of the body ( Figure 2 View Figure 2 (a, b)). Ciliated creeping sole extending to about 50% of body width. Relative position mouth:body length, of 68%. Relative position gonopore:body length, 80%.
Internal morphology
Epidermis and its secretions
Epidermis 25 µm thick. Granular coarse erythrophil and finely cyanophil secretions, respectively, are discharged through entire body surface; the former type is more abundant in dorsal and marginal epithelium, the latter, in ventral epithelium. Rhabditogen cells are densely arranged under dorsal and marginal body surface. There is no glandular margin.
Cutaneous musculature in the prepharyngeal region
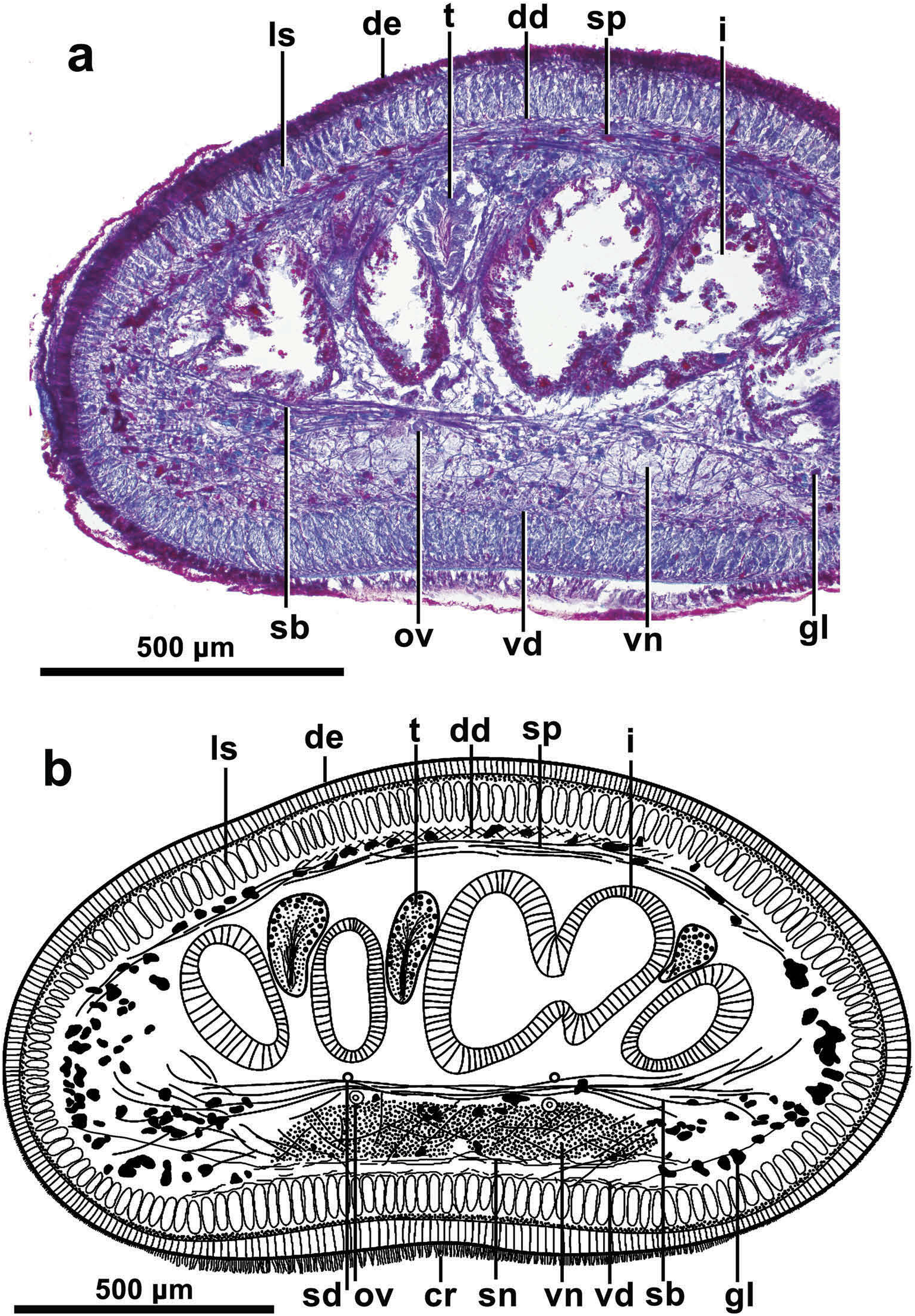
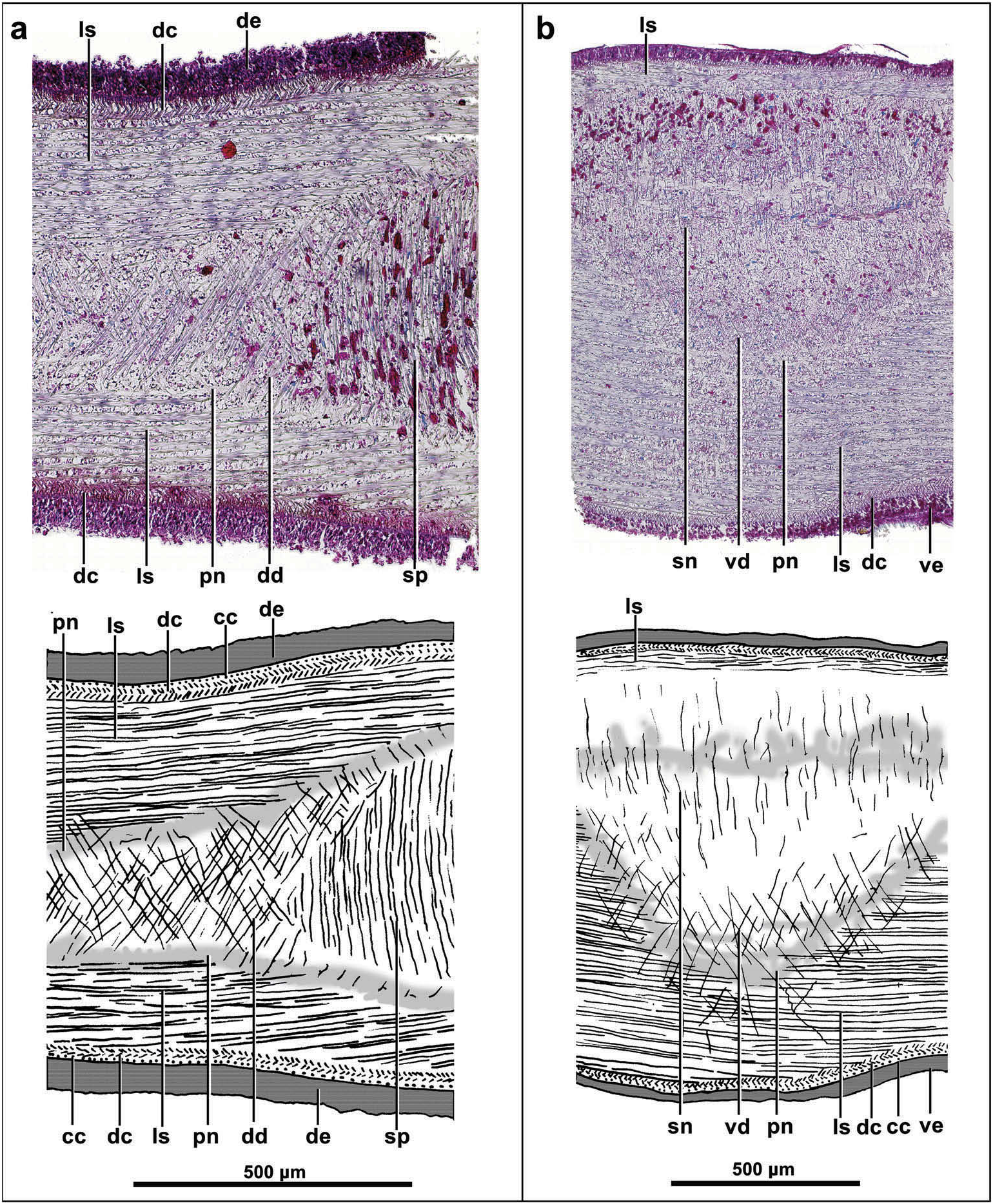
The cutaneous musculature is formed by the three typical layers found in Geoplaninae , i.e. a subepithelial one-fibre-thick circular layer, a 10 µm thick double diagonal layer with decussate fibres, and an innermost longitudinal layer; the latter layer is 75 µm thick dorsally, and 90 µm ventrally ( Figures 3 View Figure 3 , 4 View Figure 4 (a, b)). The longitudinal layer consists of dense muscle bundles. And the longitudinal layer is also well developed in the body margins ( Figure 3 View Figure 3 ). The cutaneous musculature thickness relative to body height at the prepharyngeal region is 16–19%.
Parenchymal musculature
Five parenchymal muscle layers are present throughout the body: a dorsal layer of decussate fibres, located to the inside of the peripheral nervous plexus; this layer is 10 µm thick in the mid-dorsal region, decreasing rapidly laterally until the margin, where it no longer exists; a dense supra-intestinal layer of transverse fibres (60 µm thick); a dense sub-intestinal layer of transverse fibres (60 µm thick); a subneural layer (30 µm thick) of transverse fibres; and a ventral layer (15 µm thick) of decussate fibres ( Figure 4 View Figure 4 (a, b)). Most fibres of the ventral layer lie to the inside of the peripheral nervous plexus underlying the ventral body wall; a small number of fibres of this muscle layer pierce the peripheral nervous plexus (cutaneous nervous net), and reach the longitudinal cutaneous muscle layer intersecting some of the fibres of this layer ( Figure 4 View Figure 4 (b)).
Musculature in cephalic region
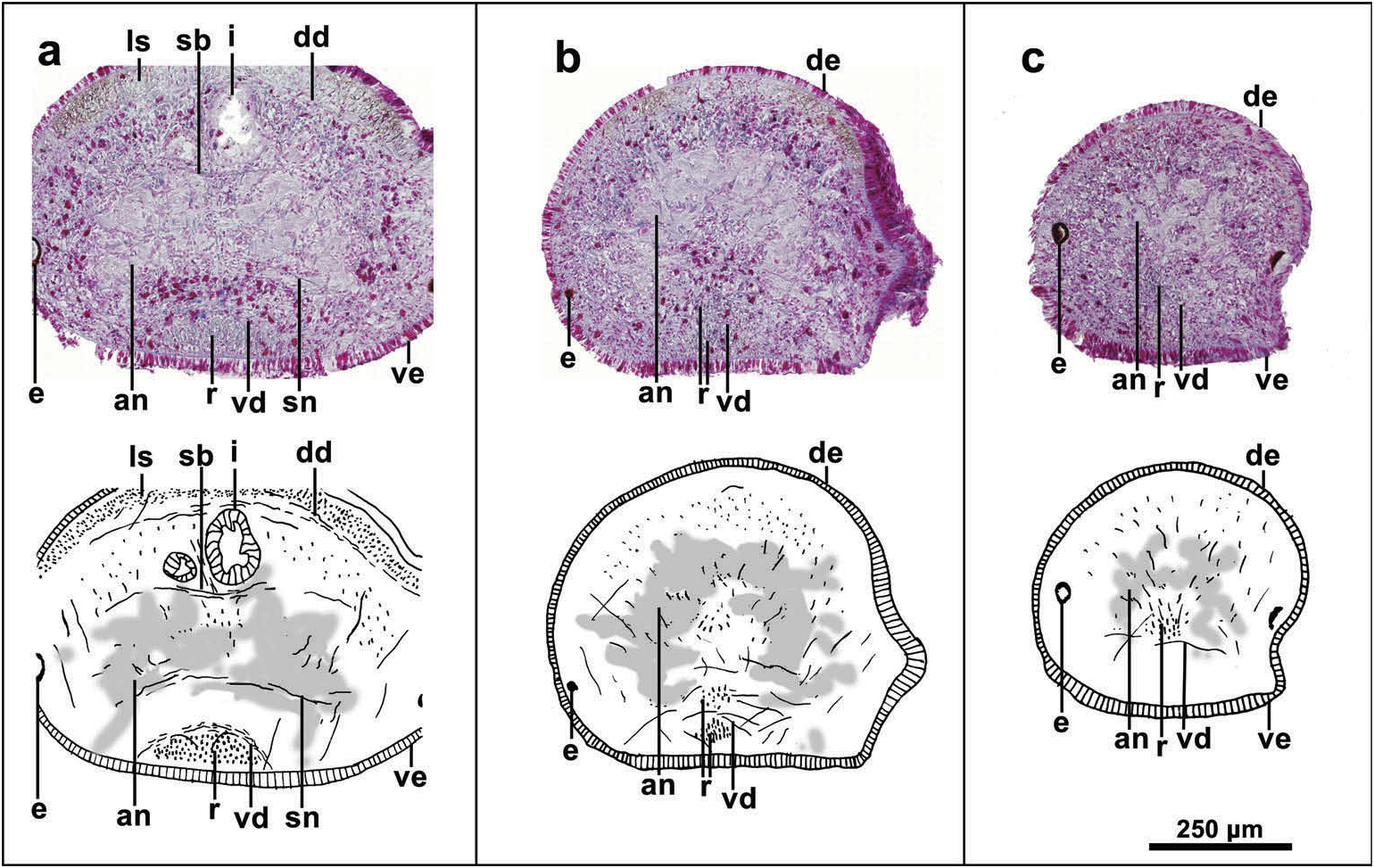
Two millimetres from the anterior tip of the body, the circular and diagonal cutaneous muscle layers are inconspicuous, while the longitudinal cutaneous layer thickens dorsally up to 100 µm and thins ventrally, reaching 75 µm. In this region, the relative thickness of the cutaneous musculature is 28%. One millimetre from the anterior tip, the creeping sole is narrower and the muscle fibres of the ventral longitudinal layer gather medially, giving rise to the cephalic retractor muscle, which is lens-shaped in cross section at this region. At 600 µm from the tip the retractor decreases in size; here, muscle fibres coming from the ventromedial region of the body run obliquely towards the dorsum and body margins, but it is unclear if these fibres derive from the retractor. At this point, the retractor is semilunate in cross section and delimited by fibres of the ventral parenchymatic muscular layer ( Figure 5 View Figure 5 (a)). At 400 µm from the tip, the retractor muscle is divided into an outer and inner bundle of fibres, both irregular in shape in cross section ( Figure 5 View Figure 5 (b)). At this point, only the ventral parenchymatic layer is developed and consists of scattered fibres. Oblique muscle fibres are abundant dorsally to the cerebral ganglia, but details of their orientation were not assessed. At 100 µm from the anterior tip, the creeping sole is lacking and the retractor muscle sinks completely into the parenchyma before its fibres disperse obliquely towards the body margins and dorsal side ( Figure 5 View Figure 5 (c)).
Digestive system
The mouth is located at a distance from the anterior portion of pharyngeal pouch equivalent to 66% of the length of the pouch ( Figure 6 View Figure 6 ). The pharynx is cylindrical in shape, and placed horizontally. It measures about 1 mm long and occupies almost 3/4 of the pharyngeal pouch. The oesophagus is 200 µm long. Both dorsal and ventral insertions of the pharynx are located approximately at the same level. The pharyngeal pouch musculature is composed of a thin outer layer of circular muscle fibres followed by an inner layer of scattered muscle longitudinal fibres. The outer pharynx musculature is composed of a layer (5 µm) of sub-epithelial longitudinal fibres followed by a layer of circular fibres (12 µm). The inner pharynx musculature (55 µm) is composed of a thick subepithelial layer of circular fibres with intermingled longitudinal fibres.
Male reproductive system
The testes are ovoid or pear shaped, most measuring about 150 µm by 320 µm. They can be as high as 30% of the body height. They are located dorsally, beneath the transverse supraintestinal parenchymal muscle layer, and between the intestinal branches ( Figure 3 View Figure 3 (b)). The anteriormost testes are located at a distance from the anterior end, equivalent to ~40% of body length; the posteriormost testes are located 1 mm before the root of the pharynx (or at a distance equivalent to ~60% of body length).
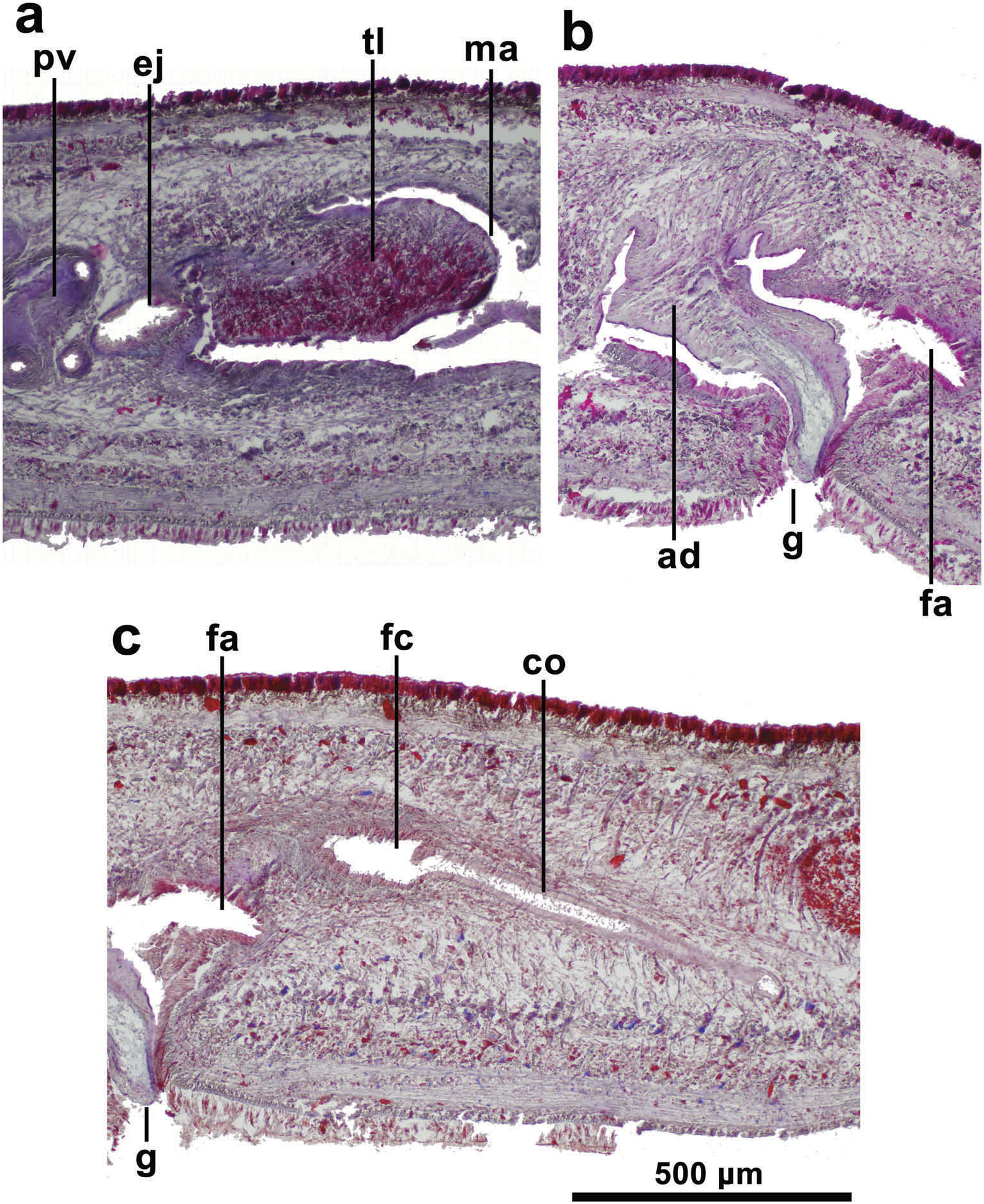
The sperm ducts run immediately above the sub-intestinal parenchymal muscle layer and slightly external to the ovovitelline ducts; they contain spermatozoa. Laterally to the proximal region of the prostatic vesicle, these ducts bend anteriorly and subsequently curve to the sagittal plane to open separately to the lateral proximal section of the prostatic vesicle ( Figure 7 View Figure 7 ). The prostatic vesicle, extrabulbar, is tubular, very long, and repeatedly recurved. Distal to the prostatic vesicle the ejaculatory duct enlarges rapidly to form an intrabulbar sub-ovoid ejaculatory cavity ( Figures 7 View Figure 7 , 8 View Figure 8 (a)). The prostatic vesicle is lined with a squamous-to-cuboidal, ciliated epithelium. It is pierced by glands producing coarse erythrophil granules and surrounded by a very dense circular musculature about 60 µm thick, distally decreasing to 10 µm. The proximal portion of the prostatic vesicle and its muscularis are immersed in a mass of scattered muscle fibres, becoming more numerous distally to finally merge with those surrounding the male section of the copulatory apparatus. The penis bulb comprises a moderate number of muscle fibres, part of which penetrates the basis of the penis papilla-like fold.
The ejaculatory cavity opens at the tip of an inconspicuous, horizontal penis papilla-like projection of the anterior wall of the male atrium ( Figures 7 View Figure 7 , 8 View Figure 8 (a)). The ejaculatory cavity is lined with a cuboidal-to-columnar ciliated epithelium and pierced by glands producing erythrophil granules. The external wall of the ejaculatory cavity consists of a 10 µm-high circular muscle layer. The male atrium is a 1.1 mm long cavity. Ventrally, this atrium is devoid of folds. Dorsally, an extension of the penial bulb projects as a musculo-glandular tongue-like large fold occupying half of the atrium, and is 400 µm long and 200 µm wide ( Figures 7 View Figure 7 , 8 View Figure 8 (a)). The tongue-like fold projects from the dorsolateral left portion of the atrium towards the right side. This fold is separated from the remaining atrium by a kind of slightly muscularized lateral sheath, which is projected from near the basis of the tongue-like fold. At its base, the sheath merges with a distinct finger-shaped musculo-glandular projection or adenodactyl ( Figures 7 View Figure 7 , 8 View Figure 8 (b)). The adenodactyl is 500 µm long and projects from the roof of the male atrium ventrally and backwards into the gonopore canal and thus separates the male from the female atrium.
The inner cavity of the male atrium is lined with a squamous epithelium, becoming cuboidal in its posteroventral region. Regarding the inner folds housed in the male atrium, i.e. the tongue-like fold, the lateral sheath, as well as the adenodactyl, they are lined by squamous epithelium. Although the penis papilla-like fold epithelium is partially damaged, it appears also to be squamous. The atrial epithelium is pierced by glands producing erythrophil granules; while the epithelium of the tongue-like fold is pierced by very numerous glands producing strongly amorphous, erythrophil secretion and some glands producing cyanophil amorphous secretion. Additional muscle fibres from the penis bulb penetrate the stroma of the anterior portion of the tongue-like fold. The male atrium is wrapped with subepithelial circular muscle layer around 10–15 µm thick, followed by a 15 µm thick longitudinal muscle layer. The squamous epithelium of the adenodactyl displays the nuclei immersed in the underlying muscular layer and is followed inwards by a thin circular muscle layer and an innermost longitudinal muscle layer. The stroma of the adenodactyl is filled by weakly cyanophil granules.
Female reproductive organs
Ovaries not found in the sections. They remain unsectioned in a body portion posterior to the cephalic and anterior to the pre-prepharyngeal section. Vitellaria poorly developed. The ovovitelline ducts run posteriorly above the central nervous plate and directly underneath the transverse sub-intestinal parenchymal muscle layer ( Figure 3 View Figure 3 ). The final portion of these ducts was not found in the sections except for the very distal one, which turns dorsomedially to enter the 600-µm long common ovovitelline duct running dorsoanteriorly at a 25° angle. It communicates with the 300-µm long female genital canal, which is a dilated, nearly horizontal projection from the posterior wall of the female atrium ( Figures 7 View Figure 7 , 8 View Figure 8 (c)). The common ovovitelline duct is lined with a cuboidal, cilated epithelium and is surrounded by a 3 µm thick, seeming circular, muscle layer. The female genital canal is lined with a cuboidal-to-columnar epithelium; the inner lining of the female atrium is columnar. The epithelia of both sections are pierced by glands producing erythrophil secretions, these being more abundant in the female atrium, and surrounded by a dense 20-µm thick muscular layer, consisting of, apparently, circular fibres; distally with some scattered longitudinal muscle fibres. No shell glands were observed, maybe because of the suboptimal quality of the sections.



| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |