
Pezothrips Karny, 1907
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4743.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:3096E7FB-4886-4410-8E46-211D07FB01B3 |
|
DOI |
https://doi.org/10.5281/zenodo.3687910 |
|
persistent identifier |
https://treatment.plazi.org/id/D529D00E-FF8D-FFC5-FF01-D0DAF6908D6C |
|
treatment provided by |
Plazi |
|
scientific name |
Pezothrips Karny |
| status |
|
Pezothrips Karny, 1907: 45 . Type species Physopus frontalis Uzel, 1895 , by monotypy.
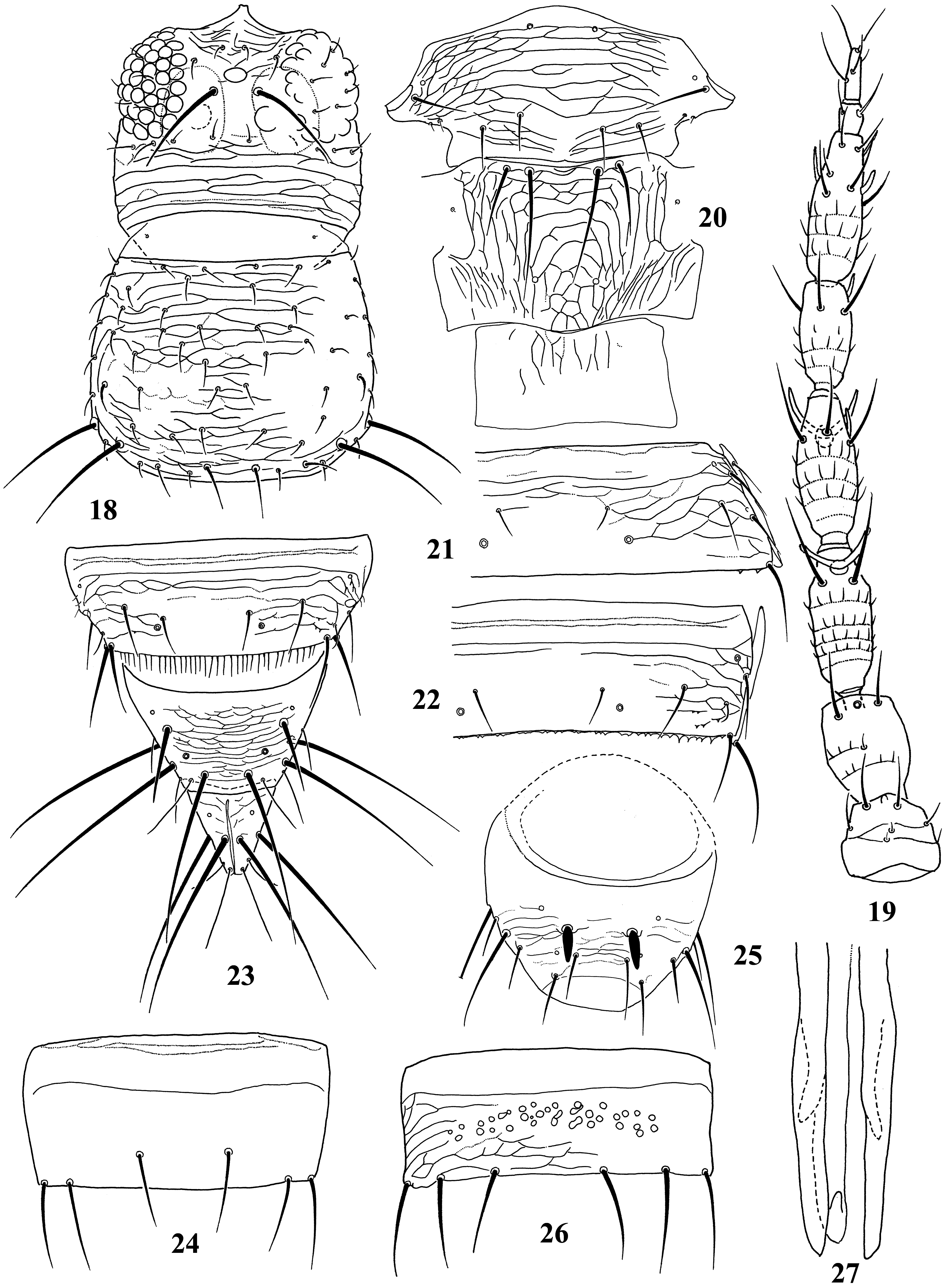
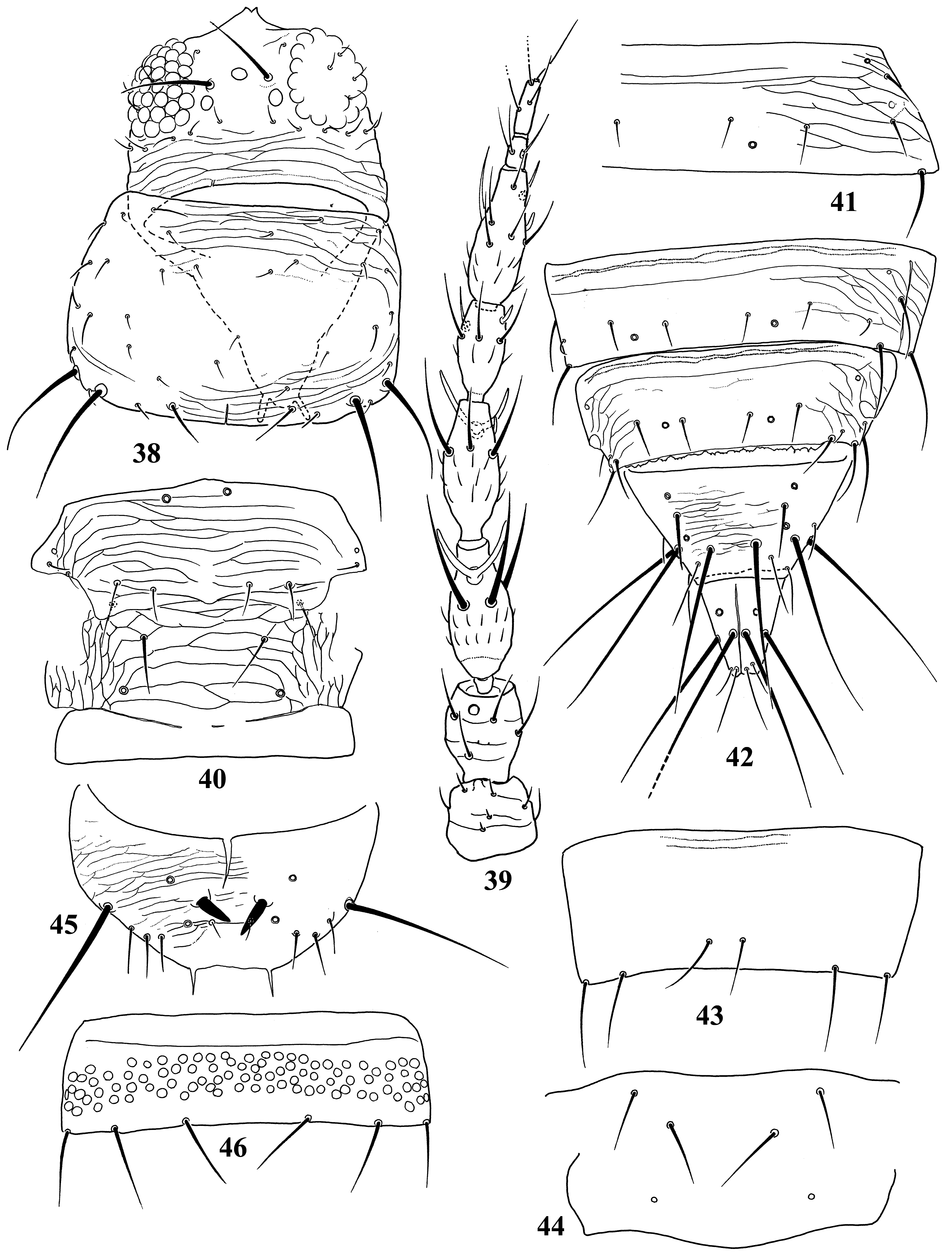
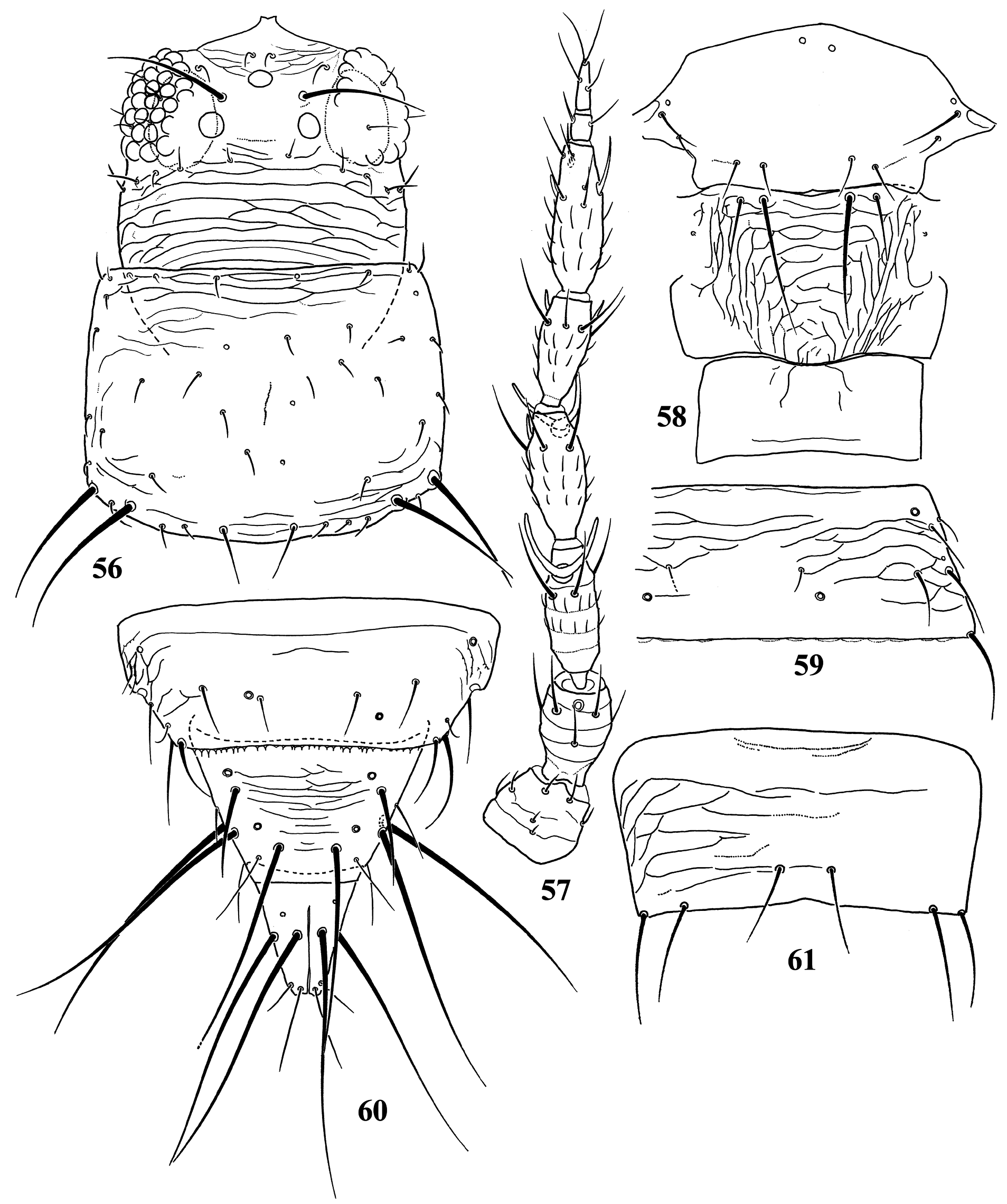
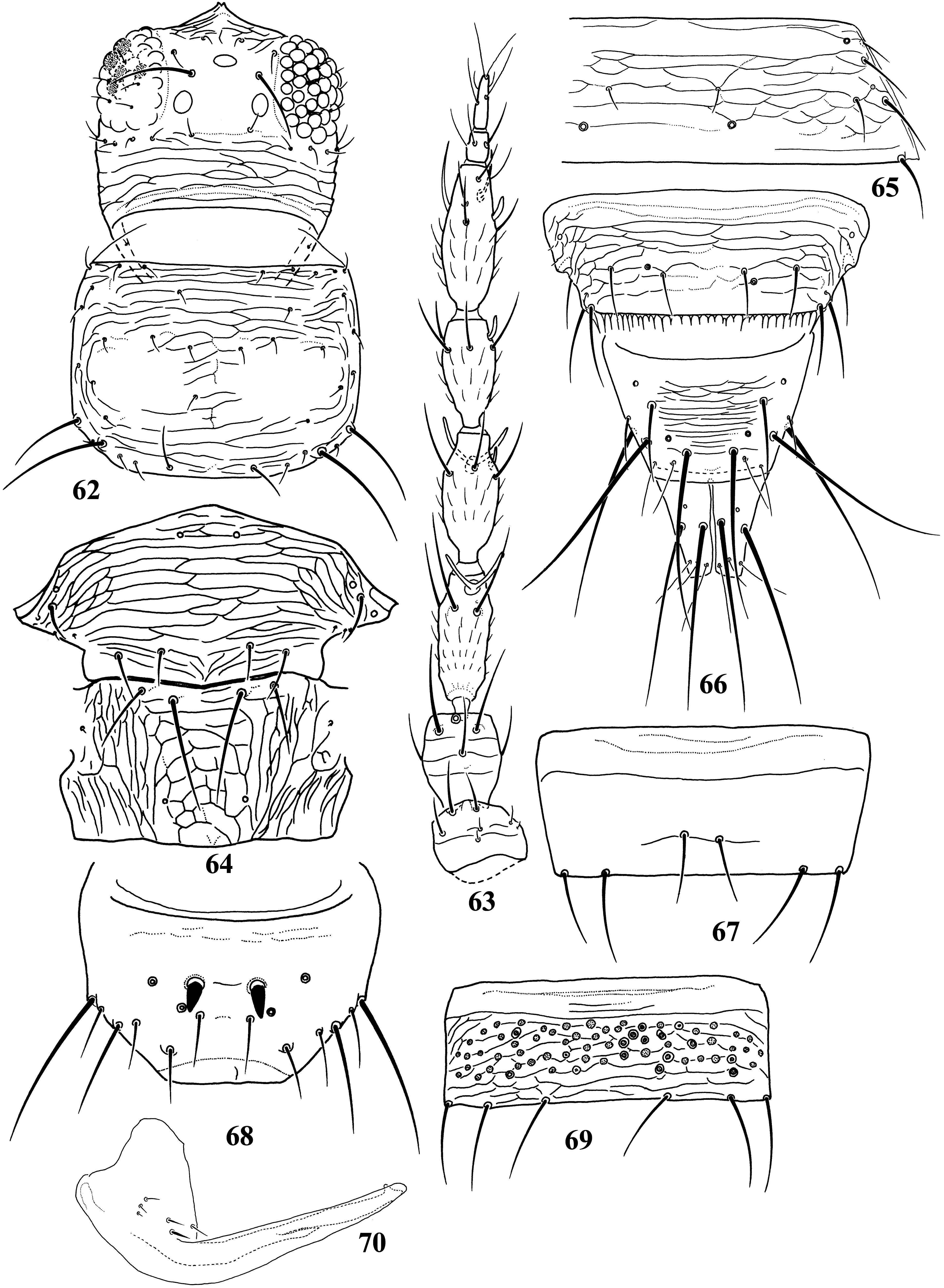
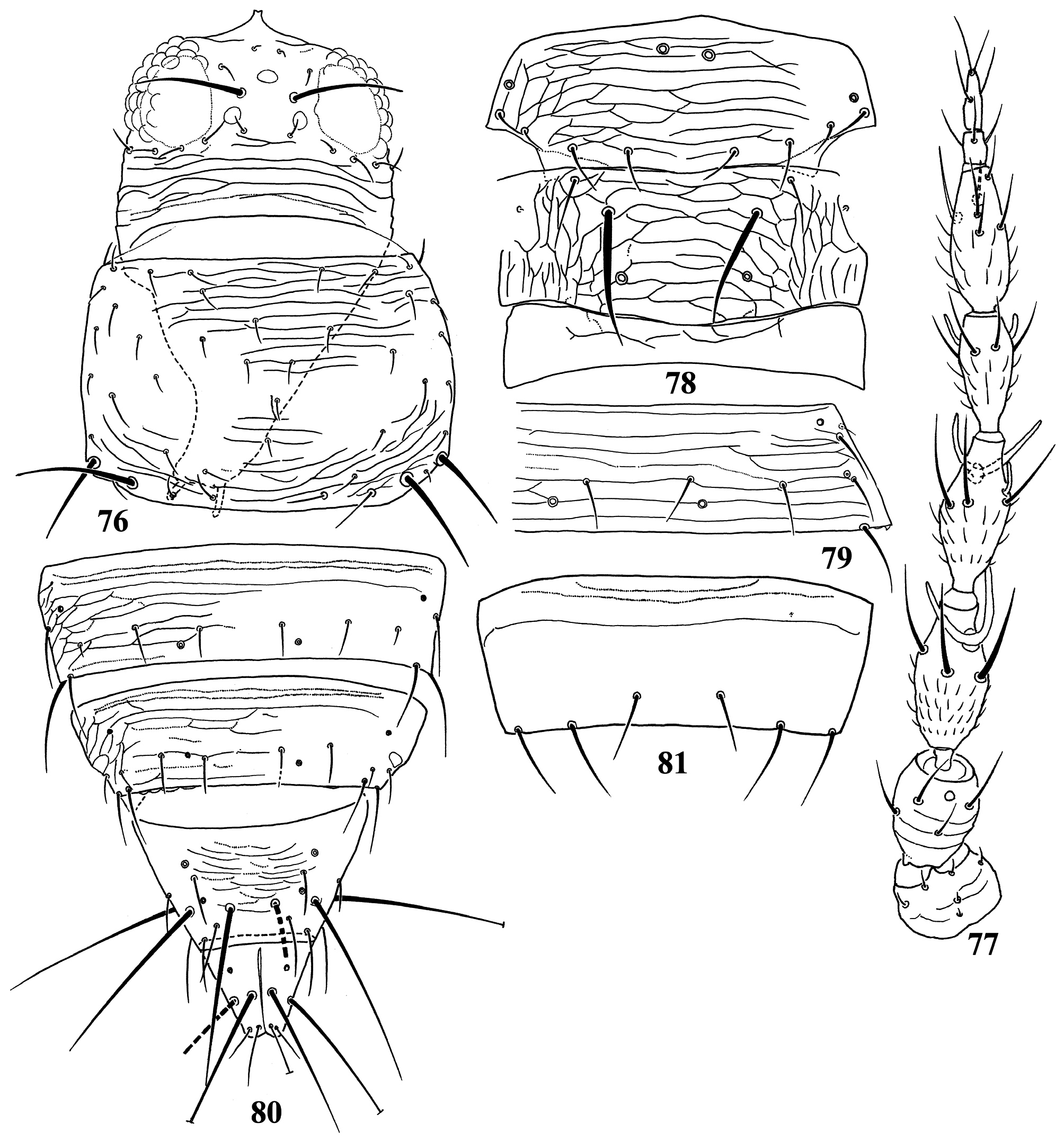
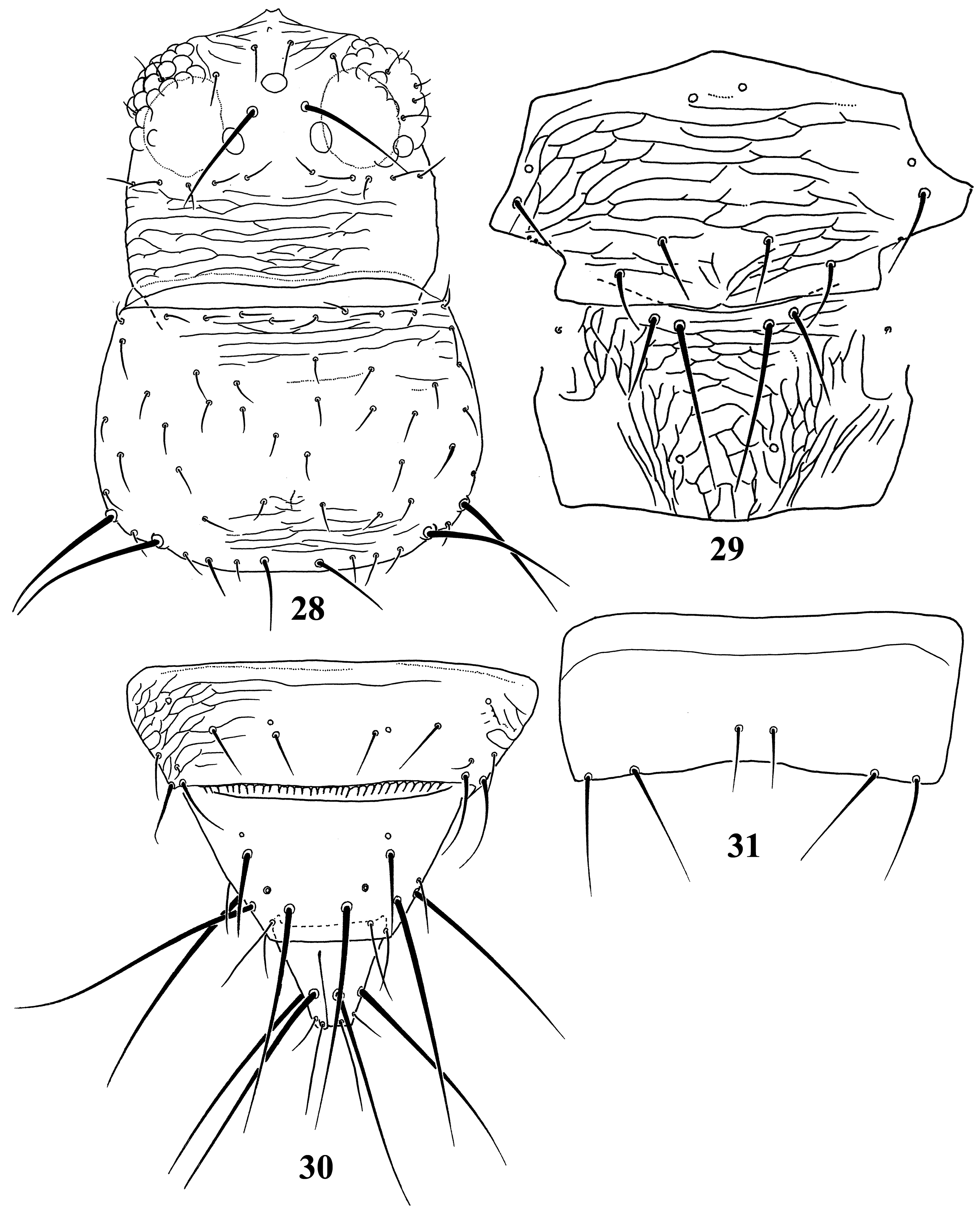
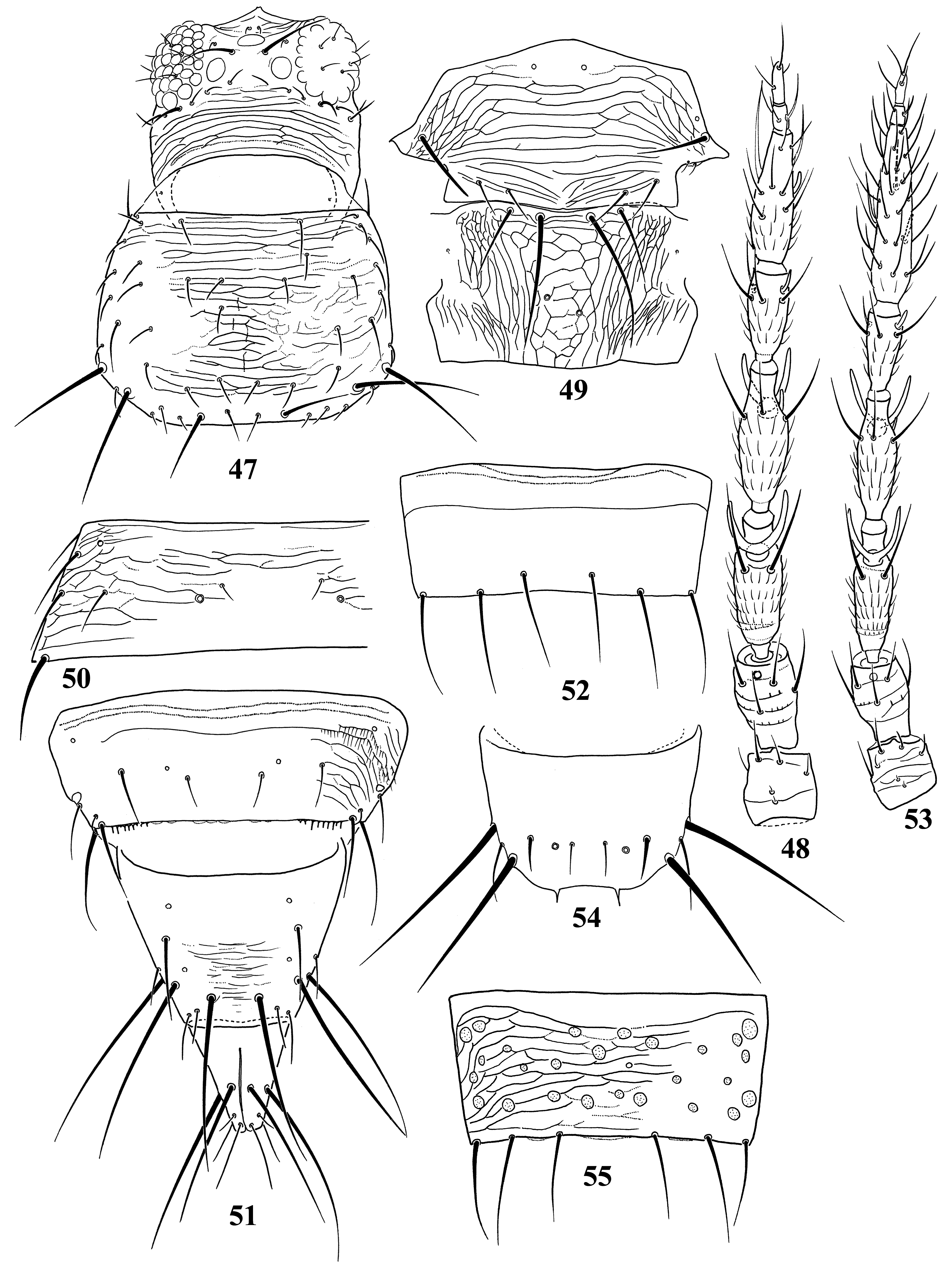
Diagnosis. Wings usually fully developed. Ocellar setae I usually present, setae III developed ( Fig. 18 View FIGURES 18–27 ). Postocular setae 5 pairs. Antennae 8-segmented, segment I with dorso-apical setae (absent in kaszabi ), III and IV with forked sense-cones, III–VI with microtrichia both on dorsal and ventral surfaces ( Fig. 19 View FIGURES 18–27 ). Pronotum with 2 pairs of posteroangular setae. Metascutum with median pair of setae at anterior margin, but often behind anterior margin in microptera. Endofurca with spinula at mesosternum only. Fore wing, when fully developed, first vein with long gap in setal row, 7–9 basal and 2 distal setae, second vein with many setae evenly spaced; clavus with 5 marginal and 1 discal setae. Abdominal tergites without ctenidia or posteromarginal craspeda; tergite II with 3 lateral marginal setae and a small seta near anterior margin on pleurotergite, and usually with a small pore anterior to S4 setae (second seta from posterior angle of tergite) ( Figs 21 View FIGURES 18–27 , 41 View FIGURES 38–46 , 59 View FIGURES 56–61 , 65 View FIGURES 62–70 , 73 View FIGURES 71–75 , 79 View FIGURES 76–81 ); tergite VIII with or without posteromarginal comb ( Figs 23 View FIGURES 18–27 , 42 View FIGURES 38–46 ); tergite IX with MD setae developed; tergite X with median longitudinal split; sternites without discal setae; sternite VII with S1 setae in front of posterior margin and usually close to each other, S2 and S3 setae at posterior margin ( Figs 31 View FIGURES 28–31 , 75 View FIGURES 71–75 ). Male with abdominal tergite IX having a pair of short, stout setae medially (except for kellyanus ) ( Fig. 36 View FIGURES 32–37 ), sternites III–VII each with many scattered pore plates ( Figs 26 View FIGURES 18–27 , 37 View FIGURES 32–37 , 46 View FIGURES 38–46 , 55 View FIGURES 47–55 , 69 View FIGURES 62–70 ); parameres each with a ventral spine at middle, hypomere with a small dorsal tubercle near apex (except for kellyanus ) ( Figs 27 View FIGURES 18–27 , 70 View FIGURES 62–70 ).
Comments. Mound & Palmer (1981) recognized Megalurothrips and related genera as Megalurothrips genus-group with Ceratothripoides , Ceratothrips , Craspedothrips , Lefroyothrips , Odontothripiella , Odontothrips and Projectothrips , but Ceratothrips , Lefroyothrips and Projectothrips are not closely related to this group because of lacking dorso-apical setae on antennal segment I. Pezothrips shares many character states with Ceratothripoides , for example fore wing first vein with long gap in setal row and male sternites with many scattered pore plates, but Ceratothripoides often has the clavus without a discal seta, and female sternite VII with S1 and S2 setae in front of the posterior margin ( Mound & Nickle 2009). However, Zhang et al. (2019) indicated that the genera with dorsoapical setae on antennal segment I, Craspedothrips , Megalurothrips and Odontothrips , are included in the same clade and this clade was sister-group to Mycterothrips , but not to Ceratothripoides in phylogenetic analysis based on morphological data. Moreover, in their analysis, Ceratothripoides seems to be sister group of Pezothrips but their systematic positions are unsolved.
The European monotypic genus, Theilopedothrips , is similar to Pezothrips but is distinguished as follows: dorsal setae on abdominal tergites not reduced, even in macropterae, male abdominal tergite IX without stout setae, sternites without pore plates, and parameres and hypomere simple ( Bhatti 1995). Asphodelothrips croceicollis is also similar to micropterous Pezothrips frontalis but has no dorso-apical setae on antennal segment I, male tergite IX has 2 pairs of stout setae, and the pore plate on male sternites III–VII is transverse, not scattered ( Bhatti 1995; zur Strassen 1995, 2003).
Taeniothrips kaszabi was transferred to Pezothrips by Bhatti (1995) for having female abdominal sternite VII with setae S1 close to each other, although from the figure with the original description it appears to have antennal segment I without dorso-apical setae ( Pelikan 1972). P. kellyanus differs from other members of the genus in having abdominal tergite VIII with paired clusters of many microtrichia, female sternite VII with S1 setae apart from each other, and male tergite IX with median pairs of setae slender. Moreover, male of Megalurothrips species generally has no sternal pore plates but Prof. Tsutsumi of Fukushima University, Japan indicated numerous lanceolate discal setae on male sternites of Megalurothrips distalis that may be associated with sternal glands (Tsutsumi, personal communication), and males of M. sjostedti have a single minute glandular opening on some sternites ( Krueger et al. 2015). Thus, kellyanus seems to belong to Megalurothrips in which it has been included (Mound & Walker 1981; Palmer 1987) rather than in Pezothrips . However, Megalurothrips species have only a short gap in the setal row on the fore wing first vein and breed in Fabaceae flowers, whereas P. kellyanus is associated with flowers of Citrus and Myoporum insulare (Webster et al. 2006) . Nguyen et al. (2016) indicated that P. kellyanus originated from Australia because mitochondrial and nuclear genetic diversity in Australia is greater than in Europe and Mediterranean region. Thus, P. kellyanus may be congeneric with neither Megalurothrips nor Pezothrips .
The distribution areas of Ceratothripoides and Pezothrips are separate as follows; species of Ceratothripoides are generally distributed in Africa, although C. claratris is from southeast Asia, whereas Pezothrips consists of the Palaearctic species indicated above. Moreover, Retanathrips , which shares several character states with these two genera, is known only in the New World ( Mound & Nickle 2009). This situation of distribution pattern seems to be similar to relationship among some genera of Megalurothrips genus-group, Megalurothrips , Odontothrips and Odontothripiella , which are associated Fabaceae flower. Megalurothrips is generally distributed in southeast Asia, although a few species are known from Ethiopian and Palaearctic regions, but Odontothrips and Odontothripiella are distributed in the Holarctic region and Australia, respectively ( Pitkin, 1972a, b). However, commonality of host plants is not found in Ceratothripoides , Pezothrips and Retanathrips .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |