
Pheretima subanensis, Aspe, Nonillon M. & James, Samuel W., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3881.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:FE9048E9-DE3A-4502-A95E-27EE8F706AC3 |
|
DOI |
https://doi.org/10.5281/zenodo.5670442 |
|
persistent identifier |
https://treatment.plazi.org/id/5B458787-FFA1-FFB0-FF5A-FE52E481BDEB |
|
treatment provided by |
Plazi |
|
scientific name |
Pheretima subanensis |
| status |
sp. nov. |
Pheretima subanensis n. sp.
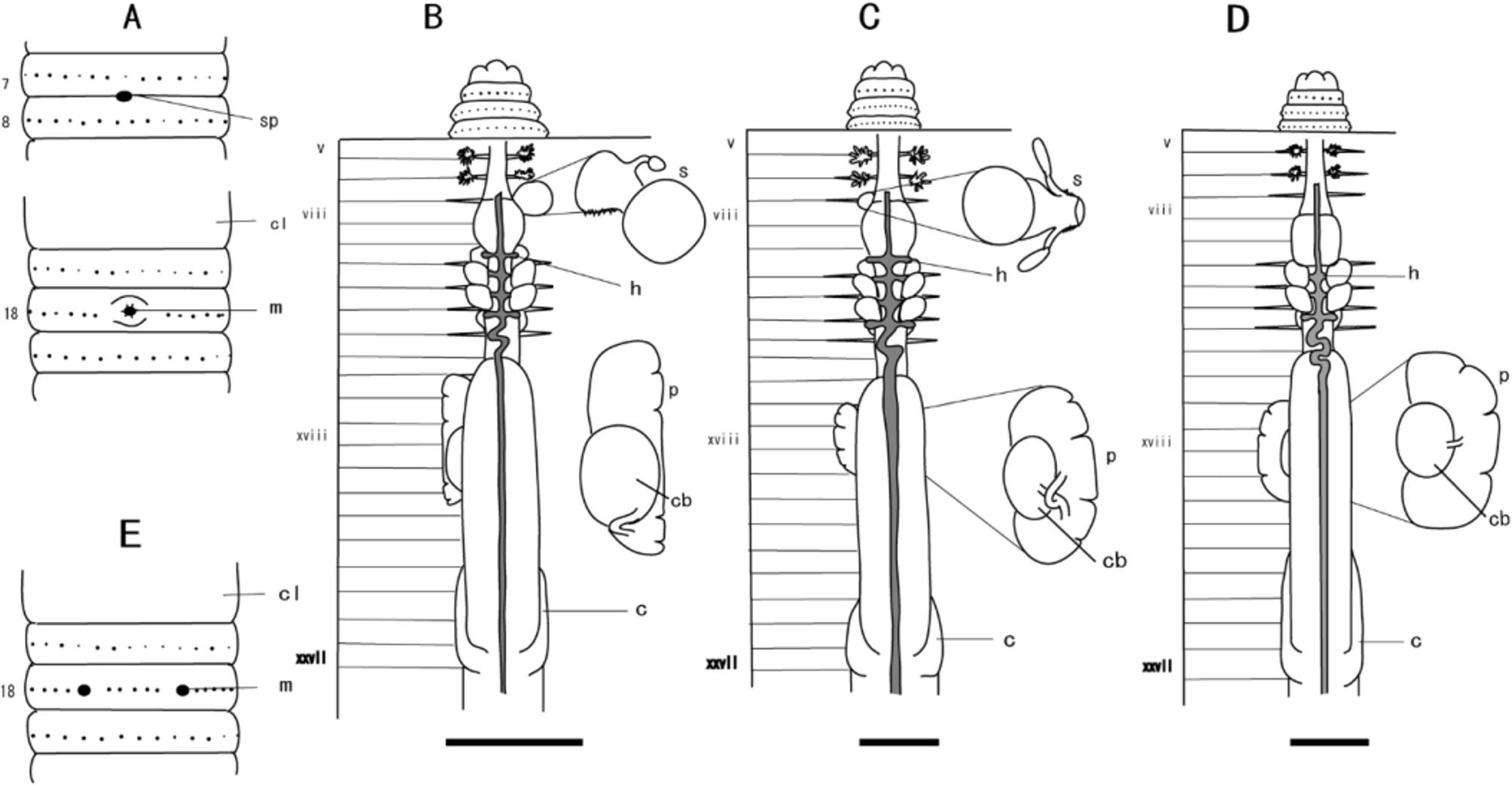
( Fig. 8 View FIGURE 8 D, Table 4)
Material examined. Holotype: adult, NMA 4523 Brgy Sibucal, Oroqieta City, Misamis Occidental Province, Mt. Malindang Range (8º19'31"N, 123º38'02"E), 991 m asl., Mindanao Island, Philippines, coll. Nonillon Aspe, M. Lluch, and J. Adeva, Feb. 18–25, 2004. Paratype: adult, amputee ( ZRC.ANN.0024), same collection data as above.
Etymology. The species is named after the indigenous tribe of Mt. Malindang, the “Subanen".
Diagnosis. Small, purplish brown worm reaching around 67 mm in adult length; ventral gap absent; first dorsal pore at 12/13; hearts paired in xi to xiii, absent in x; spermathecae absent; racemose prostate glands in xvii to xx, with columnar copulatory bursae in xviii to xix; penes lacking; caeca in xxvii to xxiii.
Description. In living animals, dorsum dark purplish brown anteriorly, fading to medium brown posteriorly; equators pigmented; clitellum off-white; ventral side non-pigmented. Length 67 mm (holotype); diameter 3.5 mm at x; 3.5 mm at xx; body cylindrical in cross-section, tail blunt; 119 segments. First dorsal pore at 12/13; spermathecal pores absent; female pore single in xiv; openings of copulatory bursae paired on xviii, 0.21 circumference apart ventrally, 5 setae between openings. Clitellum brown, annular, extending from xiv to xvi. Setae unevenly distributed, 36 setae on vii, 45 setae on xx, dorsal gap present, ventral gap absent.
Septa 5/6/ 7/8, 10/11–13/14 thinly muscular, 8/9/10 absent. Dense tufts of nephridia on anterior faces of 5/6 and 6/7; nephridia of intestinal segments located mainly on body wall at anterior and posterior faces of septa, at septum/body wall junction. Large gizzard in ix to x; esophagus with circumferential lamellae extending from xi to xii, with digitiform internal texture from xiii to xiv; intestine originates in xv; caeca originate in xxvii, extend forward to xxiii; typhlosole a simple fold of about 1/4 lumen diameter, originating at 26/27.
Hearts in xi to xiii, esophageal; commissural vessels in vi, vii, ix lateral; those in viii extend to gizzard; supraesophageal vessel extends from x to xiii; extra-esophageal vessels join ventral esophageal wall in x, receive efferent parieto-esophageal vessels in xiv.
Ovaries and funnels free in xiii; spermathecae absent. Male sexual system holandric; testes and funnels appear reduced compared to other species, enclosed in paired ventral sacs in x and xi; seminal vesicles in xi and xii, each with short dorsal lobe; vasa deferentia slender, free from body wall, passing around base of copulatory bursae en route to ental end of prostatic ducts; prostates in xvii to xx, each racemose, 3-lobed. Ductlet from anterior prostatic lobes joins vas deferens and posterior ductlet at common junction with muscular prostatic duct. Copulatory bursae in xviii to xix columnar, with rounded apex, attached to body wall at end; oriented towards common opening through body wall. Coelomic surface of bursae muscular, secretory diverticula lacking; penes absent.
Remarks. Sims & Easton (1972) stated that it was not possible for them to distinguish between athecate forms of Pheretima and Metaphire Sims & Easton, 1972 provided that the only character that distinguishes the two genera is the presence of nephridia on the spermathecal ducts in Pheretima and the absence of nephridia in that position in Metaphire . The two genera are identical in other characters such as the origin of the intestinal caeca in xxvii and the presence of copulatory bursae. Sims & Easton (1972) noted that several species assigned to Metaphire might belong in Pheretima sensu stricto, and vice versa, since the presence or absence of nephridia on the spermathecal ducts was often not mentioned in descriptions. By the same token, generic assignment to Pheretima or Metaphire is problematic for athecal worms, because spermathecal ducts are lacking, making the presence or absence of nephridia on the ducts a moot point.
Here, we propose that the relative size of the copulatory bursae may be a distinguishing character between Pheretima and Metaphire , with species in Pheretima (e.g., this study) tending to have more prominent domeshaped, intra-coelomic copulatory bursae than those in Metaphire (e.g., Ohfuchi 1938, 1957; Tsai et al. 2004; Bantaowong et al. 2011), and if this is the case, it would be useful in assigning athecal worms to one or the other of these genera ( Chang et al. 2009). Homology among copulatory bursae has not been clear; some authors consider only invaginations of the body wall into the coelom to comprise copulatory bursae, whereas others also consider intramural chambers and even shallow indentations to be copulatory bursae. In the case of Pheretima vs. Metaphire as defined by Sims & Easton (1972), the type of invagination is the same in the two genera: a large or small bursa visible from within the body, and this is the source of the difficulty in assigning athecal worms in the Pheretima complex with caeca and copulatory bursae to one or the other of these genera. The criterion of having a large or small, invaginated copulatory bursa would seem to exclude from either genus any species whose male pore invaginations are entirely within the body wall (intramural), or whose male pores lie in shallow indentations.
Examples of species originally identified as Pheretima but reassigned to Metaphire by Sims & Easton (1972) because nephridia on the spermathecal ducts were not mentioned in descriptions are members of the octothecal, holandric M. ignobilis Gates, 1935 species group, which are most similar to P. darnleiensis . Members of this group apparently have only intramural male pores but not intra-coelomic copulatory bursae as in true Pheretima . One member of this group, M. riukiuensis Ohfuchi, 1957 , has only thick epidermal lips pressed together covering the male pores, rather than an invagination. None of these species would be considered synonyms of P. darnleiensis , and it is clear that a major taxonomic revision of Metaphire is necessary.
Island Locality N [species]
Luzon 30 Kalbaryo 4
Kalinga 6
Mt. Province & Banaue1 9
Mt. Arayat 2
Mt. Makiling 2
Mt. Isarog 7
Catanduanes 2
Mindanao 34 Mt. Kitanglad 15 Mt. Malindang 18 Mt. Apo2 1
Cebu 1
Total 67
1 Includes nominal P. darnleiensis , also reported from Papua New Guinea, Fiji, Malaysia, Indonesia, and Singapore. 2 Includes P. urceolata , also reported from Indonesia.
Joshi et al. (1999) reported an athecal earthworm 200–300 mm long from Ifugao, Luzon, but it is likewise not clear whether this species belongs in Pheretima or in Metaphire . James (unpublished) however thought that this species is most likely a Pheretima basing on the size and shape of the copulatory bursae. Aspe (unpublished data) also described another athecal species> 300 mm long at Bukidnon, central Mindanao, with prominent domeshaped copulatory bursae, and result on molecular analysis shows that this species belongs to the Pheretima clade rather than that of Metaphire . Pheretima subanensis n. sp. is similar to P. malindangensis n. sp., P. vergrandis n. sp., and P. potonganensis n. sp. in relative size and coloration, but it is athecal, whereas the latter three are bithecal, monothecal and octothecal, respectively. The athecal species also differs from the others in male pore spacing, setal gaps, the size and position of the prostate glands and copulatory bursae, and the intestinal origin in xv rather than in xvi.
Gates (1972) listed the following as indicators that lumbricid or megascolecid earthworms are parthenogenetic: (1) testes and/or seminal vesicles retained in the juvenile state in adult specimens; (2) absence of spermatozoal iridescence in male funnels and/or spermathecae; (3) spermatophores lacking or, if present, lack spermatozoa inside. In Pheretima subanesis n. sp., the testes appear to be reduced, and we observed no spermatozooidal iridescence in the sperm funnels. These observations suggest that this species is parthenogenetic.
Occurrence. Pheretima subanensis n. sp. was found in disturbed forest in Brgy Small Potongan, at 915–1024 m elevation (Table 1).
| ZRC |
Zoological Reference Collection, National University of Singapore |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Pheretima |