
Petaloconchus varians, (D' ORBIGNY 1841)
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00058.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B6B923-EE3B-FFED-8F86-66C23AF06F7B |
|
treatment provided by |
Carolina |
|
scientific name |
Petaloconchus varians |
| status |
|
PETALOCONCHUS VARIANS (D’ ORBIGNY 1841) View in CoL
Material Examined
Sebastian Inlet, Florida ( USNM 890935).
External anatomy and mantle cavity
Operculum thin and chitinous, shallowly concave, slightly overlapping side of foot. Spiral lamella upstanding for approximately one half volution, forming raised ridge for another one half volution. Single, low, inconspicuous spiral lamina present within shell along columellar surface. Lamina becoming obsolete towards aperture. Body of animal long and slender, appearing as sideways displaced, sometimes irregular, coils. Posterior end of body thinning to long, slender tail comprising only gonadial tissue. Columellar muscle thick anteriorly, thinning to narrow strip running almost complete length of body, unattached to viscera for much of that length. Ctenidial filaments narrowly triangular. Ctenidium extending from mantle edge, not reaching base of mantle cavity. Voluminous branchial vein traversing base of mantle cavity from pericardium to ctenidium. Hypobranchial gland well developed, composed of transverse folds covering rectum and mantle roof from anus to base of mantle cavity. Cephalic tentacles short and thick, lying on either side of broad, flattened head. Two long, slender pedal tentacles lying just under snout, atop reduced foot sole. Pedal gland opening medially under pedal tentacles. Pedal gland large, extending approximately onehalf length of cephalic haemocoel, alongside oesophagus. Numerous small, ciliated ducts penetrating secretory portion of gland, coalescing into large ciliated duct under circum-oesophageal nerve ring. Pedal gland duct extending anteriorly along floor of columellar muscle to aperture.
Reproductive system
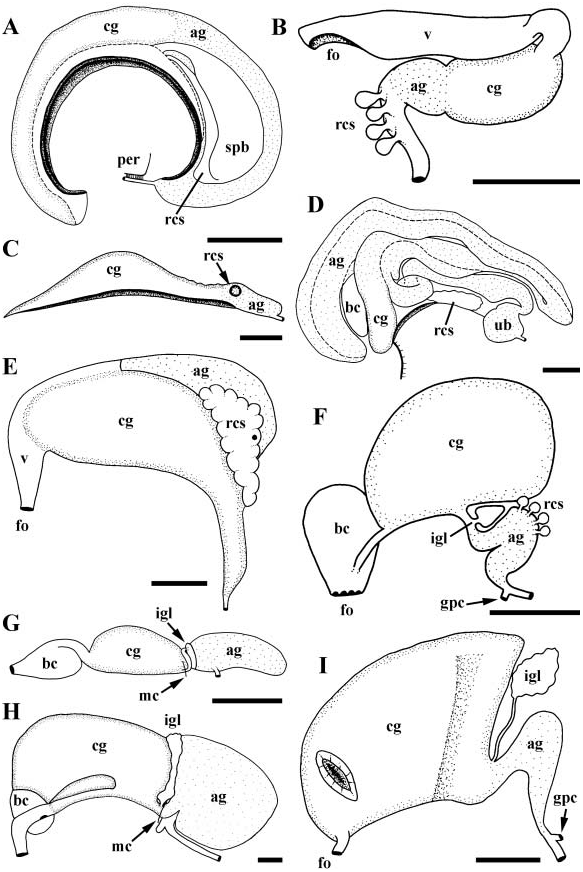
Gonad displaced posteriorly, comprising long posterior tip of visceral mass. Gonad thinning anteriorly, running along left, ventral aspect of digestive gland. Narrow oviduct emerging from ovary at posterior end of midgut, opening to pallial oviduct at base of mantle cavity. Gonopericardial duct absent. Pallial oviduct thin, short, weakly glandular, traversing basal onethird of elongate mantle cavity ( Fig. 2C View Figure 2 ). Short albumen gland (ag), approximately one-fourth size of capsule gland (cg). Single, ovoid seminal receptacle (rcs) present at anterior end of albumen gland. Oviductal groove closed along proximal albumen gland, opening at level of receptacle. In brooding females, anterior one-third of mantle roof split along midline, accommodating attachment of egg capsules to interior of shell. Egg capsules, attached via narrow thread short distance back from shell aperture, containing numerous small embryos. Attached sacs not arranged in linear series of oldest to youngest, but occurring in mixed groups.
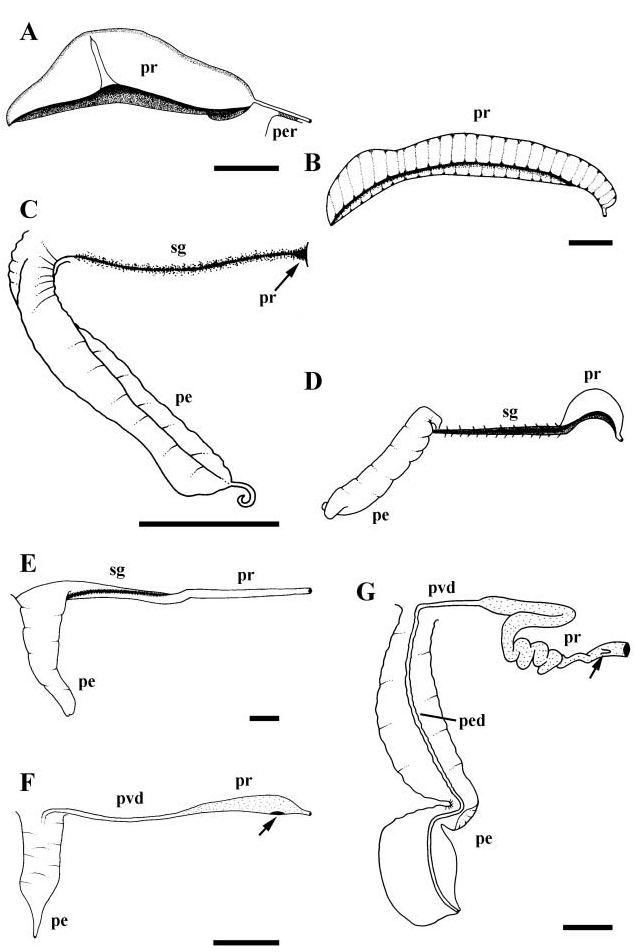
Vas deferens emerging from testis at posterior end of gastric chamber, widening rapidly, engorged with ripe sperm, functioning as seminal vesicle. Seminal vesicle narrowing toward base of mantle cavity and opening to pallial gonoduct ( Fig. 3B View Figure 3 , pr). Prostate composed of two highly glandular laminae with deep intervening groove, extending approximately one-half length of mantle cavity. Laminae largely open, fused only short distance along proximal end. Laminae flaring widely, exposing glandular surfaces to mantle cavity.
Alimentary system
Foregut. Radula taenioglossate. Paired jaws present at anterior ends of dorsal folds; jaw composed of rods with laterally overlapping homogeneous layer. Sub-radular membrane incompletely covering odontophore. Sub-radular organ present, composed of vertical, glandular crest projecting into mouth opening. Salivary glands passing through circumoesophageal nerve ring. Radular sac short, curving dorsally around right posterior limit of buccal cavity. Glandular mid-ventral fold present in posterior buccal cavity. Two, short ventro-lateral folds present within anterior oesophagus. Septate oesophageal gland absent.
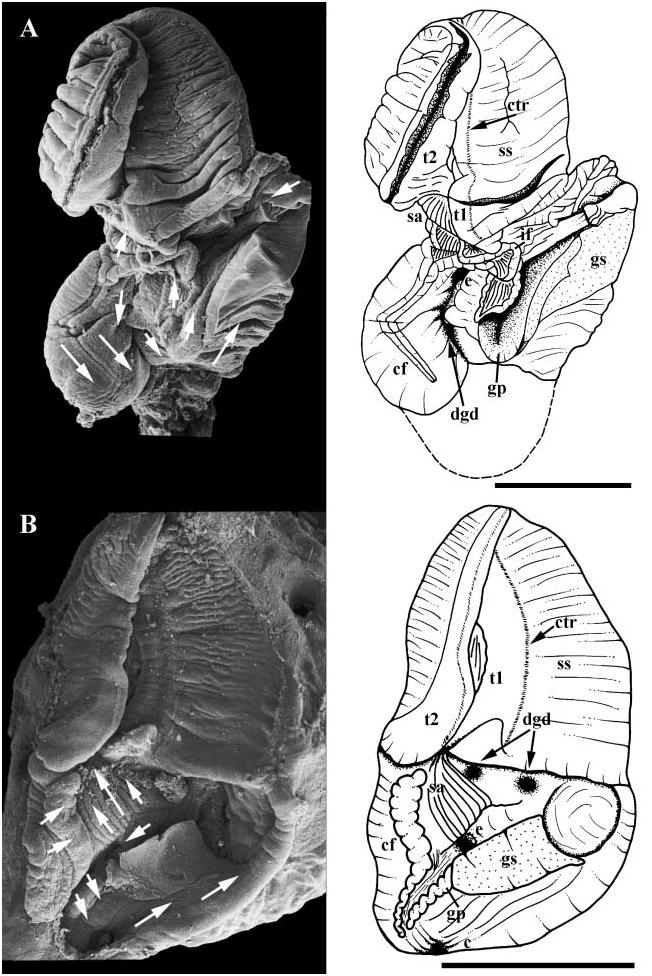
Midgut. Oesophagus opening ventrally, on left ( Fig. 14A, e View Figure 14 ). Prominent gastric shield (gs) on right, ventro-lateral wall. Well-developed sorting area (sa) extending posteriorly from intestinal groove, straddling oesophageal aperture. Sorting area covering surface of tall, free-standing flap (if) between oesophageal aperture and intestinal groove. Ciliated fold (cf) bounding sorting area and extending posteriorly from sorting area, along left wall of gastric chamber. Ciliary currents flowing toward intestinal groove within sorting area. Single digestive gland duct (dgd) present mid-ventrally. Glandular pad (gp) projecting only short distance past tip of gastric shield. Digestive gland largely resticted posterior to gastric chamber; anterior digestive gland lobe possibly absent. Style sac with paired typhlosoles and transversely folded epithelium bearing differentiated cilia. Crystalline style and raised ciliary tract (ctr) along major typhlosole (t1) present. Raised tract thin, only several cilia thick. Ciliary currents in style sac flowing counter-clockwise when viewed from behind.
Hindgut. Intestine emerging from style sac, curving dorsally to right over posterior end of kidney.
Reno-pericardial system
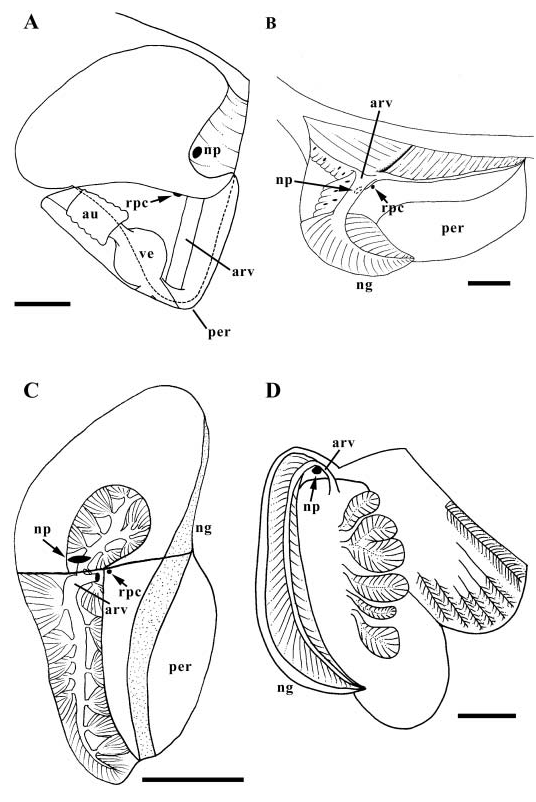
Kidney lying alongside pericardium, partially overhanging pericardial lumen, extending into pallial roof. Pallial and visceral portions of kidney approximately equal in size ( Fig. 22C View Figure 22 ). Afferent renal vessel entering kidney just behind base of mantle cavity, extending dorsally, branching into two large vessels supplying pallial and visceral excretory tubules in roof. Renopericardial canal (rpc) opening ventrally, to left of afferent renal vessel. Region of connective tissue distinct from remaining excretory tissue (ng) present along length of kidney roof. Efferent nephridial vein joining pallial portion of efferent branchial vein.
Nervous system and sensory structures
Nervous system epiathroid, right and left dialyneurous. Nerve ring lying short distance behind buccal mass. Supra-oesophageal connective long; sub-oesophageal connective very short. Buccal ganglia lying at base of buccal mass, anterior to circum-oesophageal nerve ring. Tentacular nerve bifid. Visceral connectives extremely long, extending along columellar muscle to single visceral ganglion at base of mantle cavity. Small statocysts with single statoliths present on postero-lateral surfaces of pedal ganglia. Eyes small, present at lateral bases of cephalic tentacles. Osphradium comprising short, thin, ciliated ridge along anterior portion of ctenidium.
Remarks
Although Sebastian Inlet vermetids have been identified as Petaloconchus varians ( Mikkelsen, Mikkelsen & Karlen, 1995) , this identification should be viewed with caution. D’Orbigny’s type material includes two thin and chitinous operculae, but they are crushed and the presence of upstanding spiral lamellae could not be determined, nor could the presence of internal spiral laminae be assessed ( Keen, 1961). Moreover, much of the original description included features that may fall within the realm of intraspecific variation, notably, variations in colour and colour pattern of the mantle, foot and cephalic region ( d’Orbigny, 1834 – 46).
Olsson & Harbison (1953) described a new species of Petaloconchus from Florida that ranges from the Pliocene to the Recent, P. floridanus . They claimed that specimens of this species are commonly confused in museum collections for P. varians . They criticized the type material of P. varians as being too fragmentary and insufficient for purposes of identification, but paradoxically described the new species on the basis of coiling habit and shell ornament, features notoriously variable within vermetid species, with no mention of diagnostic features of the operculum, internal aspects of the shell, or of internal anatomy. Moreover, they did not cite unique features that could serve to distinguish P. floridanus from P. varians . Thus, the description for P. floridanus is equally inadequate for purposes of identification as the original type material for P. varians . Consequently, positive identification of the current material examined awaits re-description of the type material and comprehensive anatomical descriptions for both P. varians and P. floridanus .
Discussion
Given this important caveat, the specimens identified here as Petaloconchus varians possess many characters diagnostic of the genus which distinguish it from other operculate vermetid genera. These include the sideways displaced, spiral growth form, an elongate columellar muscle, spiral opercular lamellae and spiral laminae within the shell ( Carpenter, 1856; Keen, 1961; Morton, 1965). Petaloconchus varians is unique among described members of the genus because only a single, low lamina is present – the number and development of laminae in the genus are variable, but are often paired and sufficiently prominent to partially occlude the lumen of the shell ( Keen, 1961; Morton, 1965).
The reproductive anatomy of P. varians presents some unusual features. Of particular interest is the presence of a mantle slit in females and brooding of egg capsules attached to the shell. Several taxonomic revisions and comparative studies have emphasized the fact that the operculate genera, Petaloconchus and Dendropoma , lack both of these features and brood unattached egg capsules within the mantle cavity ( Morton, 1954, 1965; Keen & Morton, 1960; Keen, 1961; Hadfield, 1970; Hughes, 1978). With rare exception, egg capsules are ordered in a linear series from oldest to youngest in these genera ( Hughes, 1978). However, P. varians was found to brood egg capsules attached to the inner surface of the shell by a fine thread. Egg capsules were not arranged in a linear series, but occurred in mixed clusters. Egg capsules were also observed within the mantle cavity. However, it is unclear whether these were being brooded or were newly formed egg capsules awaiting attachment to the shell. In comparison to other vermetid genera, both Serpulorbis and Vermetus s.s. reportedly brood attached sacs and bear a deeply incised mantle ( Morton, 1951a,b,c, 1965; Hadfield, 1970; Hughes, 1978).
Morphology of the capsule gland also distinguishes Petaloconchus varians , forming a long, thin organ at the base of the mantle cavity. The anterior tip of the capsule gland lies well back of the posterior limit of the mantle slit. In Serpulorbis and Vermetus, Morton, 1965 ) speculated that the pouch-like capsule gland comes directly into contact with the lining of the shell in order to secure the embryo sacs. This does not seem to be possible in all species of Serpulorbis (e.g. Hadfield, 1970), nor in P. varians , indicating that an alternative method of egg capsule attachment should be explored in these species, possibly via the pedal gland. The presence of a receptacle occurring at the junction of the albumen and capsule glands is common within the Vermetidae ( Morton, 1951 a, 1965).
The vermetid digestive system has been described by Morton (1965) as offering no distinctive generic features. Existing descriptions, however, suggest that several midgut features vary and could be a potential source of characters within the group. For example, Yonge (1932) described the presence of a fold separating the opening of the intestine from the cavity of the midgut. As described above, this fold is similarly present in Petaloconchus varians , its surface bearing an extension of the sorting area. This fold is also present in Serpulorbis zealandicus ( Morton, 1951a; = S-shaped fold) and Dendropoma (Novastoa) lamellosa (as Dendropoma lamellosa ) ( Morton, 1951c; = strongly projecting fold). The folds in P. varians and S. zealandicus both bear sorting lamellae.
A second feature of interest is the digestive gland ducts, which vary from two ( Morton, 1951a,c) to three ( Yonge, 1932). The presence of a single digestive gland duct and the absence of digestive gland tubules anterior to the midgut in P. varians suggests that the anterior lobe may have been lost in this species. Confirmation of this character requires detailed study of the digestive gland and digestive gland ducts. One final character is ciliary current direction within the style sac region, which may flow in a clockwise ( Morton, 1951a) or counter-clockwise direction ( Yonge, 1932; present study). This feature similarly requires further investigation.
Although there are few accounts of vermetid nervous systems, they indicate that the nervous system is epiathroid and right dialyneurous; in contrast to the present description, a left zygoneury may be present ( Lacaze-Duthiers, 1860; Hadfield, 1970). Visceral ganglia may be single (Hadfield) or paired (Lacaze- Duthiers); the tentacular nerve may be single and the statocysts lacking (Hadfield).
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.