
Periclimenaeus alius, Park & Grave, 2021
|
publication ID |
https://doi.org/ 10.26107/RBZ-2021-0022 |
|
publication LSID |
lsid:zoobank.org:pub:4198F098-19D9-4C21-A7C3-05F26CA75086 |
|
persistent identifier |
https://treatment.plazi.org/id/94C9252B-BE24-4404-945D-7A3B0583D2DB |
|
taxon LSID |
lsid:zoobank.org:act:94C9252B-BE24-4404-945D-7A3B0583D2DB |
|
treatment provided by |
Diego |
|
scientific name |
Periclimenaeus alius |
| status |
sp. nov. |
Periclimenaeus alius View in CoL , new species
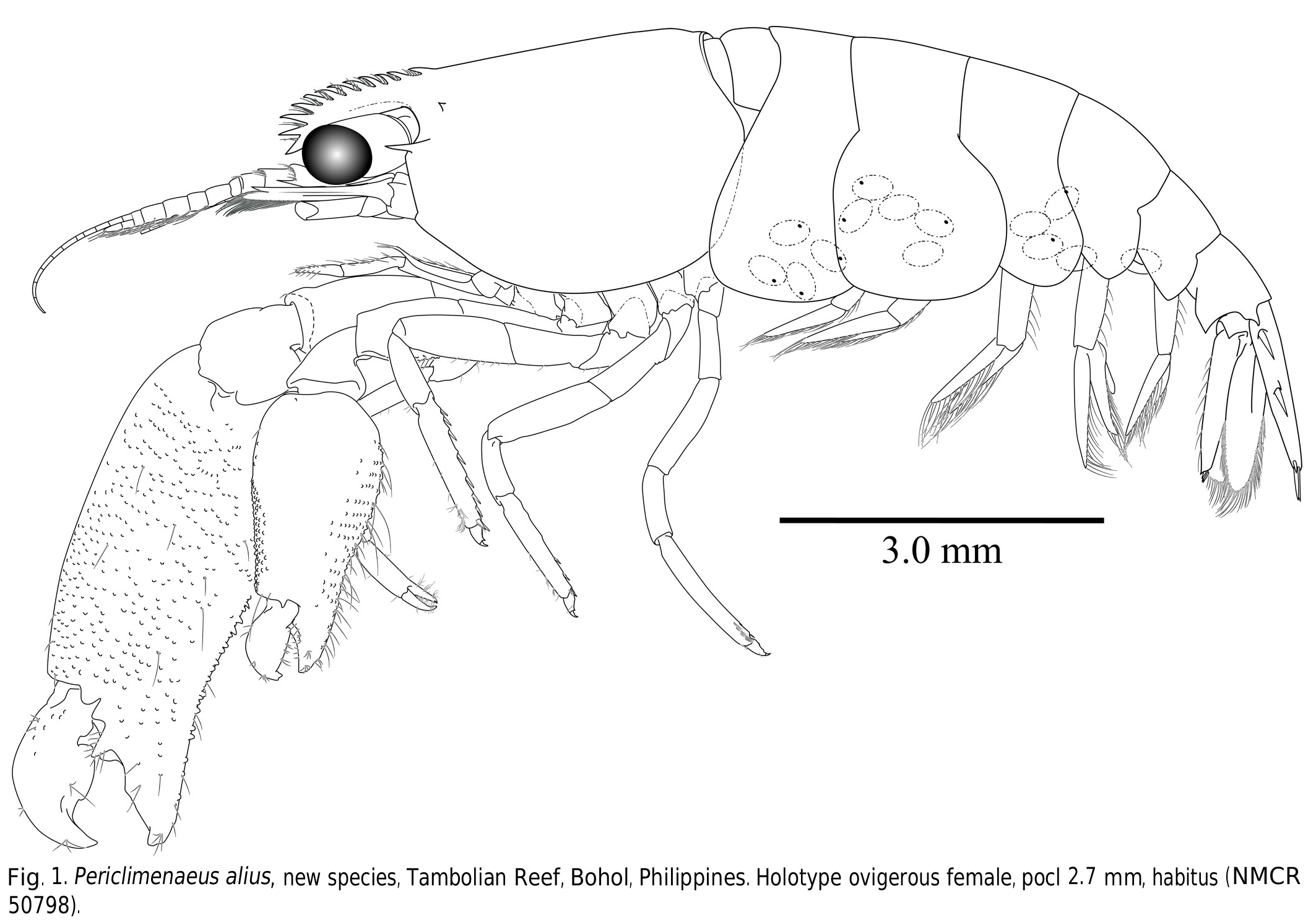
( Figs. 1–5 View Fig View Fig View Fig View Fig View Fig )
RAFFLES BULLETIN OF ZOOLOGY 2021
Material examined. Holotype: ovigerous female (pocl 2.7 mm) ( NMCR 50798 ), Tambolian Reef , Bohol, Philippines, 10 °3′20.42″N, 123°56′58.28″E, 22 m in depth, from unidentified sponge, leg. D. Lee, 9 March 2018 (fcn PB19) GoogleMaps . Paratypes: 1 male (pocl 2.8 mm) ( NMCR 50799 ), same data as holotype (fcn PB20) GoogleMaps ; 2 males (pocl 2.9, 3.1 mm) ( ZRC 2021.0382 View Materials ), same data as holotype (fcn PB18, 22) GoogleMaps .
Etymology. From the Latin adjective (nominative) ‘alius’, meaning ‘another’; alluding to the description of yet one more species in the genus.
Description (based on female holotype). Medium-sized palaemonid shrimp, body subcylindrical ( Fig. 1 View Fig ).
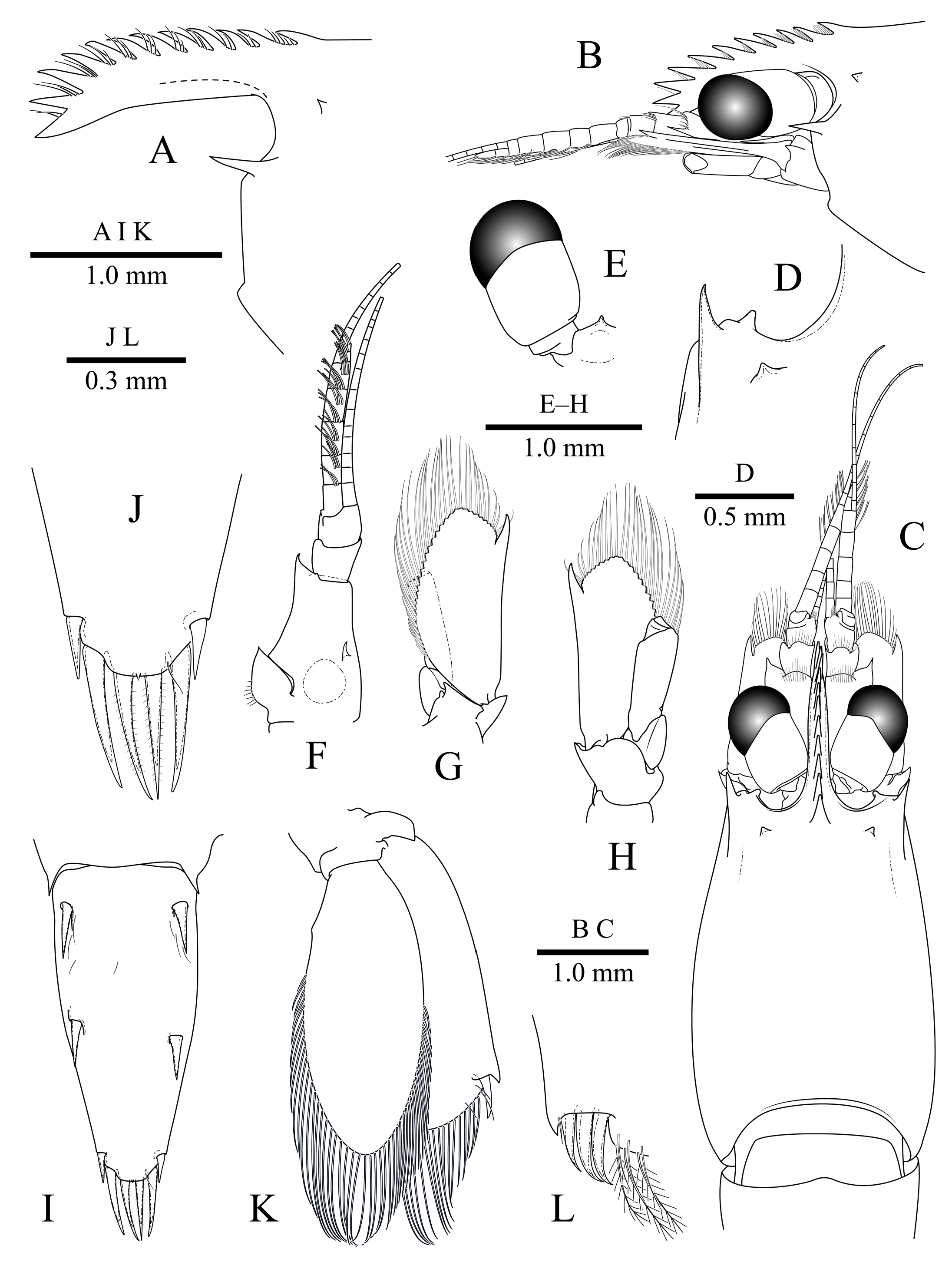
Rostrum ( Fig. 2A View Fig ) straight, trending downwards distally, about 0.45 of pocl, falling short of distal margin of antennular peduncle; furnished with 11 dorsal teeth, distally increasing in size, one of which placed post-orbitally, marginally more distant; without ventral teeth.
Carapace ( Figs. 1 View Fig , 2C View Fig ) smooth, glabrous; supra-orbital tooth feebly developed ( Fig. 2B, C View Fig ); antennal tooth acute, well developed, reaching to distal margin of basicerite; inferior orbital angle with blunt pointed process ( Fig. 2D View Fig ); pterygostomial angle produced into a sub-angular lobe. Pleon ( Fig. 1 View Fig ) smooth, first pleuron with distinct but poorly developed anteromedian dorsal lobe; first four pleura broadly rounded; fifth pleuron with posterolateral angle quadrate; sixth pleuron about same length as fifth, about 0.5 of telson length, posterolateral margin acute, median lobe acute.
Telson ( Figs. 1 View Fig , 2I View Fig ) about 0.6 of pocl, about 2.2 times longer than maximal width; with two pairs of dorsal cuspidate setae, subequal in size, inserted about 0.1 and 0.5 of telson length respectively; posterior margin ( Fig. 2J View Fig ) convex, with minute acute median process, with three pairs of spiniform setae, lateral pair short, about 0.4 of length of intermediate pair, intermediate pair longest, submedian pair about 0.9 of length of intermediate pair, setulose.
Eye ( Figs. 1 View Fig , 2E View Fig ) with hemispherical cornea, cornea faceted, about 1.6 times as long as dorsal and lateral maximal diameter, nebenauge absent.
Antennule ( Fig. 2F View Fig ) with proximal article of peduncle bearing acute distolateral tooth, ventromesial tooth present; stylocerite sharply pointed, reaching to mid-length of proximal article; intermediate article short, about 0.4 of length of proximal article, subequal to distal article; upper flagellum biramous, proximal five divisions fused, short free ramus with two divisions, longer free ramus with 11 and 12 divisions; lower flagellum filiform.
Antennal basicerite ( Fig. 2B, G, H View Fig ) with pointed process distolaterally; carpocerite reaching about 0.6 of scaphocerite; scaphocerite ovoid, about 1.7 times as long as wide, distal lamella obtusely triangular, acute distolateral tooth situated at about 0.9 of scaphocerite length, reaching to end of distal lamella.
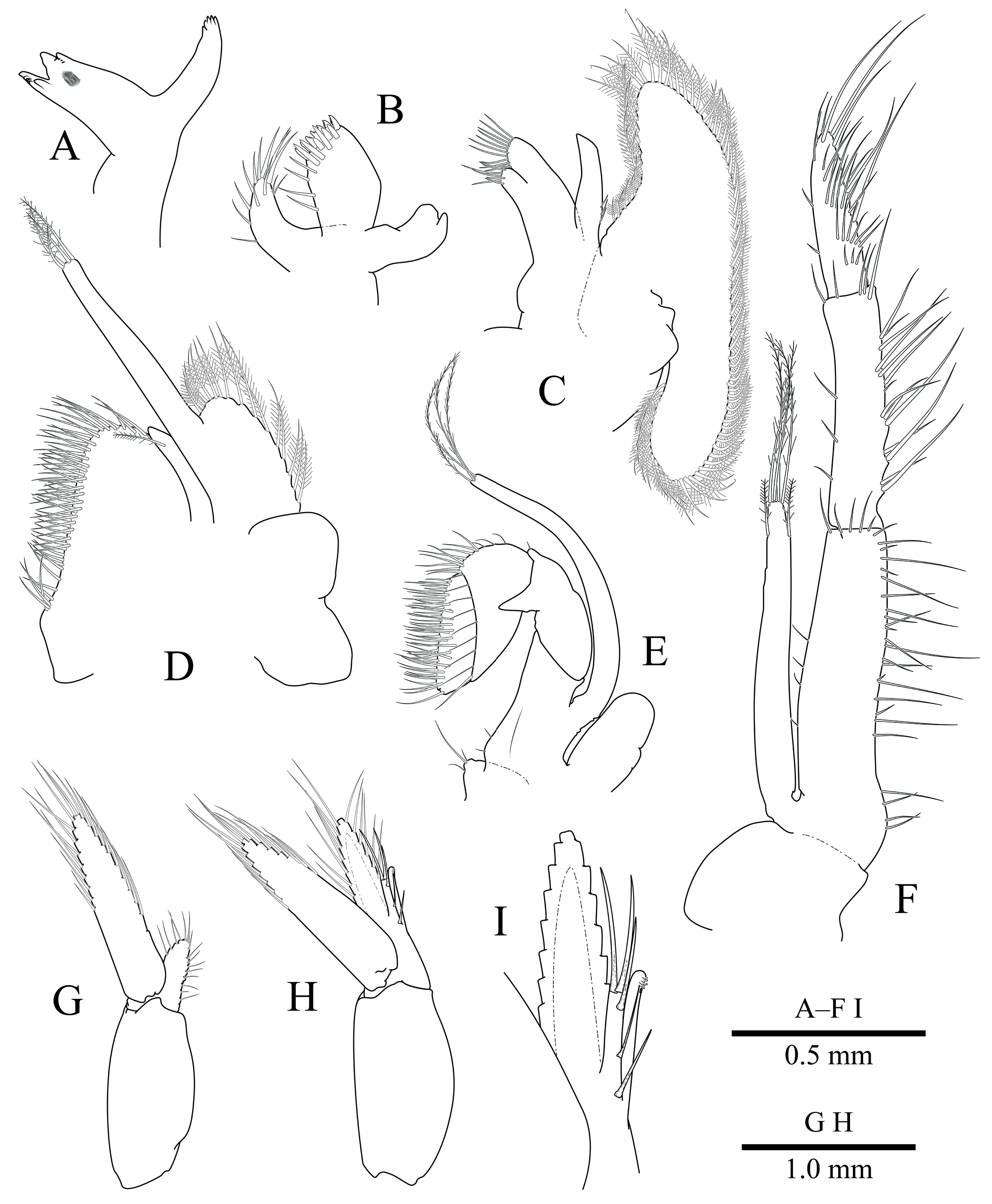
Mouthparts ( Fig. 3A–F View Fig ) as illustrated; typical for genus.
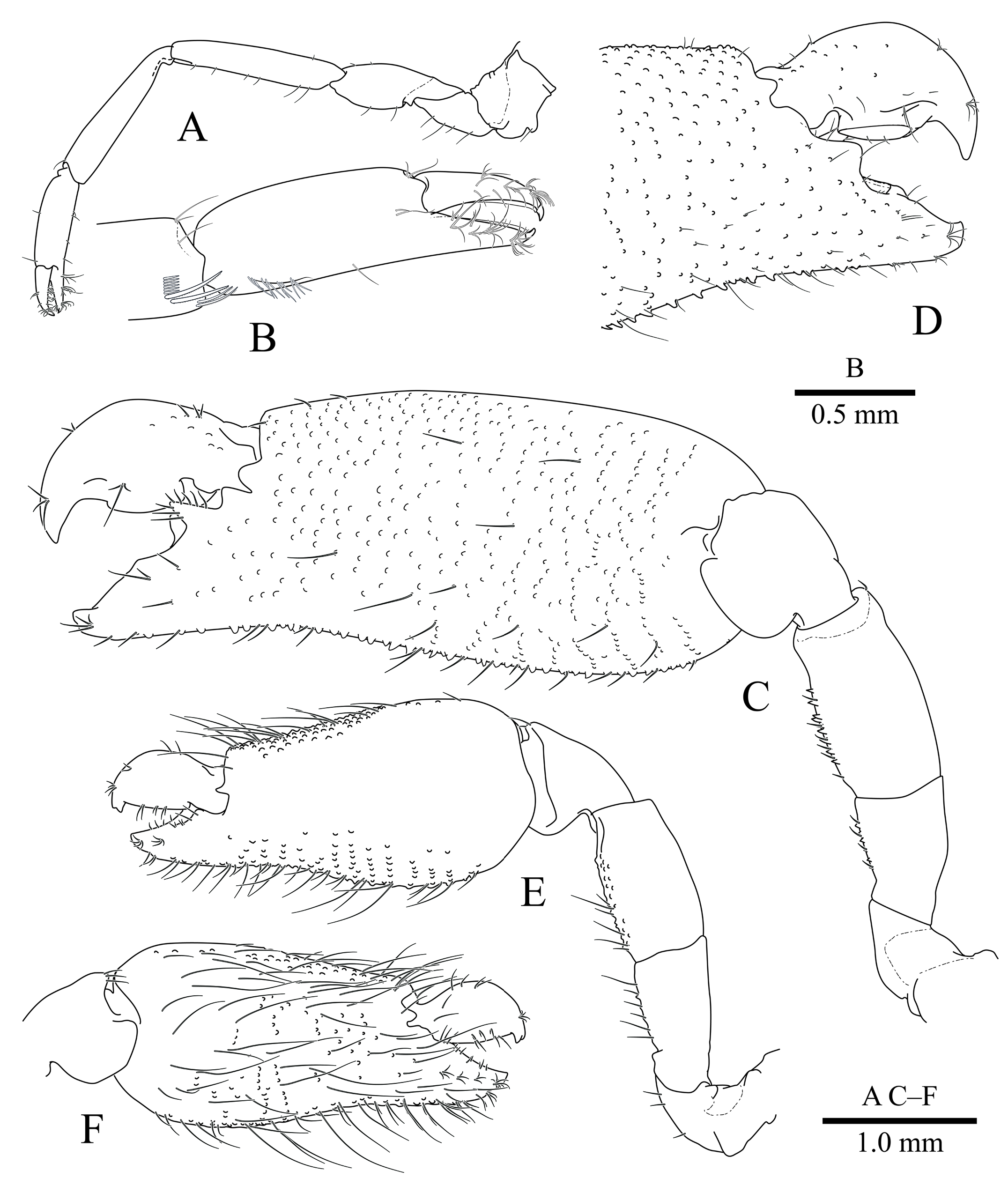
First pereiopod ( Fig. 4A View Fig ) with coxa and basis without special features; ischium about 0.75 of merus length, unadorned; merus slightly longer than carpus, unadorned; carpus subequal to chela, unadorned; chela ( Fig. 4B View Fig ) stout, palm subcylindrical, smooth, fingers about 0.5 of palm length, tips of both pollex and dactylus simple; carpo-propodal brush well developed.
Second pereiopods ( Figs. 1 View Fig , 4C, E View Fig ) robust, dissimilar in shape, unequal in size.
Major second pereiopod ( Fig. 4C, D View Fig ) with coxa and basis without special features; ischium about 0.6 of merus length, sparsely tuberculated on ventral margin; merus with conical tuberculation along ventral margin; carpus robust, about 0.8 of merus length, vase-shaped; chelae robust, about 1.85 times as long as pocl; palm subcylindrical, densely tuberculated on both margins and surfaces; fingers mesially slightly curved, about 0.24 of total chela length, pollex subequal in length to dactylus, terminating in triangularly acute tip, cutting edge entire, proximal cutting edge with triangular process on mesial margin; dactylus about 0.6 times as deep as long, cutting edge entire, distally with hooked tip, well-developed fossa and molar system present.
Minor second pereiopod ( Fig. 4E View Fig ) with coxa and basis without special features; ischium subequal to merus length, sparsely tuberculated on ventral margin; merus with fine tuberculation along ventral margin, marginally extending onto mesial and lateral surfaces, carpus short, about 0.65 times length of merus, vase-shaped; chela ( Fig. 4E, F View Fig ) robust, as long as pocl, about 0.6 of major chela length, palm subcylindrical, pronounced tuberculation along ventral and dorsal margin, extending onto adjoining lateral and mesial surfaces, central aspect of mesial surface sparsely tuberculate; fingers about 0.3 of total chela length, pollex about 0.8 of dactylus, terminating in triangularly acute tip, cutting edge entire, dactylus about 0.5 times as deep as long, cutting edge entire, distally with hooked bidentate tip.
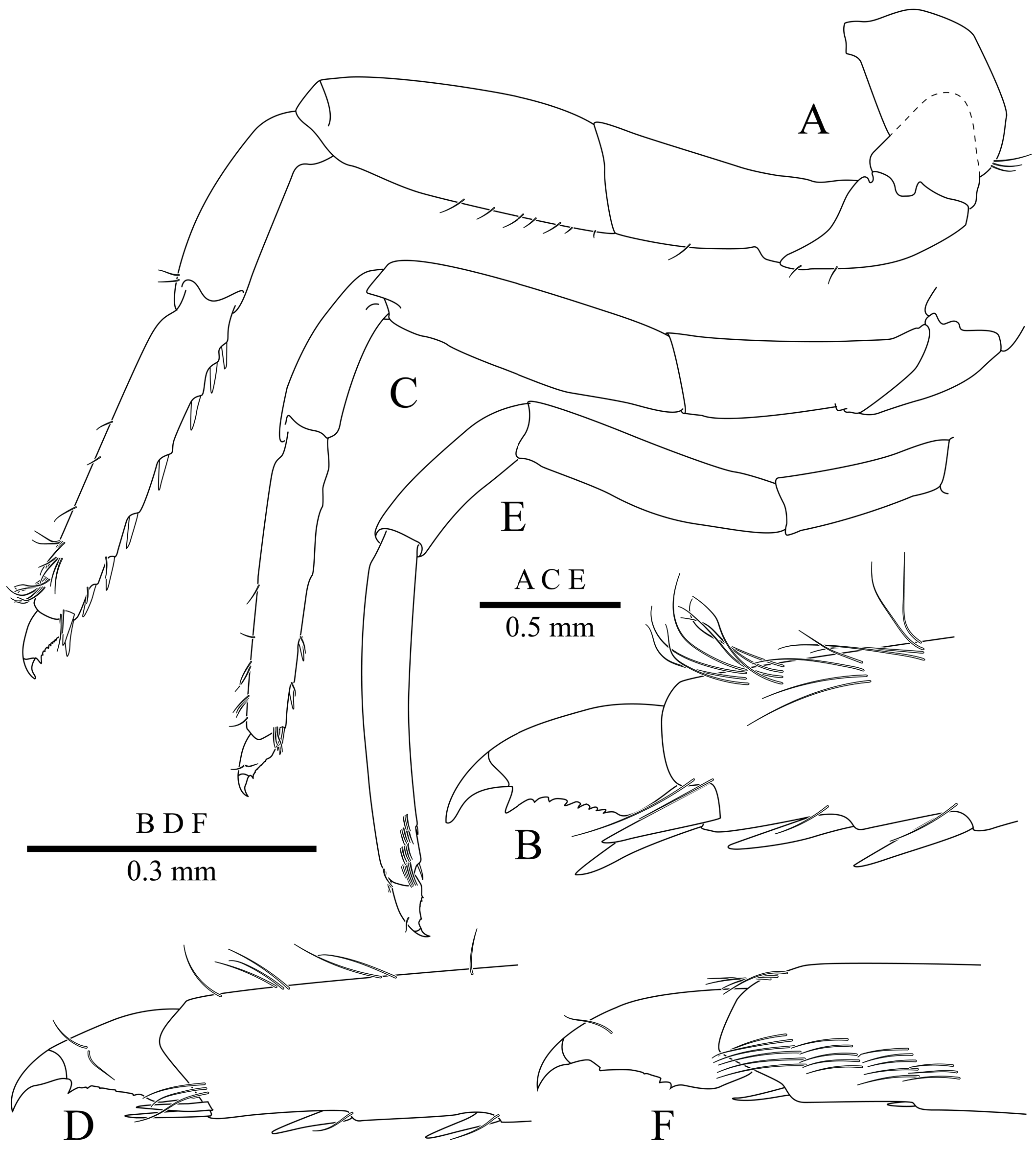
Third pereiopod ( Fig. 5A, B View Fig ) relatively robust; coxa and basis without special features; ischium about 0.8 of merus length, unadorned; merus about 1.5 of carpus length, unadorned; carpus about 0.7 of propodus length, unadorned; propodus about six times as long as maximal depth, flexor margin armed with series of single, cuspidate setae, spaced at regular intervals across entire margin, distoventral angle with pair of spiniform setae; dactylus, short, about 0.2 of propodus length, unguis demarcated, simple, curved, about 0.4 of length of corpus; corpus with distinct, distal accessory tooth, flexor margin serrated. Fourth pereiopod ( Fig. 5C, D View Fig ) generally similar to third pereiopod in proportions; ornamentation of propodus with fewer spiniform setae. Fifth pereiopod ( Fig. 5E, F View Fig ) slenderer than third or fourth, cleaning brush well developed.
Uropod ( Fig. 2K View Fig ) just overreaching distal margin of telson; exopod slightly shorter than endopod, lateral margin nonserrate, distolateral tooth well developed, adjacent spiniform seta strong (aberrant number on left uropod in holotype, Fig. 2L View Fig ), overreaching tooth.
First pleopod of male ( Fig. 3G View Fig ), as illustrated; typical for genus.
Second pleopod of male ( Fig. 3H View Fig ) with endopod with long appendix masculina in relation to appendix interna, with five long setulose setae; appendix interna about 1.4 times longer than appendix masculina ( Fig. 3I View Fig ).
Morphological variation. The general morphology of both sexes is largely identical, although the major second chela is proportionally larger in male specimens, e.g., 2.2 times in male (pocl 3.1 mm) vs. 1.85 times in the ovigerous female holotype (pocl 2.7 mm). The largest male paratype (pocl 3.1 mm) has a proportionally shorter rostrum and fewer teeth (7/0), but it is clear that the distal tip has been broken off.
Colour. Specimens preserved without colour notes or photo, no traces of colour.
Host. All specimens were obtained from an unidentified sponge.
Distribution. Currently only known from the type locality, Tambolian Reef, Bohol, the Philippines.
Remarks. A number of species groups have been proposed within the genus Periclimenaeus (e.g., robustus group cf. Bruce, 2005b; rhodope group cf. Bruce, 2006), although the majority of species have not been assigned to any groups. In the absence of a phylogeny of the genus, be it molecular or morphological, the systematic significance of these groupings and indeed the potential importance of the morphological characters on which they are based remain unclear. It therefore seems more logical to contrast the new species to other taxa within an identification framework, rather than speculating on what species could be phylogenetically closest. A clear framework for such a comparison was established by Bruce (2013), who drew attention to the utility of the third pereiopod to quickly derive at a relatively small number of potential candidate species during identification within this morphologically heterogeneous genus. Using this scheme, the new species falls within his informal Group 3, by having the dactyli of the third pereiopod with a distal accessory tooth, the corpus of which harbours ventral denticles (i.e., serrated), but with the unguis being non-dentate.
Nineteen Indo-West Pacific species are known to display this ornamentation: P. apomonosi Park & De Grave, 2021 ; P. aurantiacus Bruce, 2014d ; P. bidentatus Bruce, 1969 ; P. djiboutensis Bruce, 1969 ; P. garthi Bruce, 1976 ; P. hebedactylus Bruce, 1970 ; P. heronensis Bruce, 2010 ; P. leptodactylus Fujino & Miyake, 1968 ; P. lobiferus Bruce, 1978 ; P. matherae Bruce, 2005a ; P. mortenseni Bruce, 1994 ; P. nufu Ďuriš, Horká & Hoc, 2009 ; P. pachyspinosus Marin, 2007; P. pulitzerfinali Bruce, 2011 ; P. rastrifer Bruce, 1980 ; P. solitus Bruce & Coombes, 1995 ; P. spongicola Holthuis, 1952 ; P. tchesunovi Ďuriš, 1990 ; and P. tuamotae Bruce, 1969 .
The new species harbours a distinct, although small, supraorbital tooth, which serves to immediately distinguish it from the following species in this group in which such a tooth is absent: viz. P. djiboutensis , P. garthi , P. heronensis , P. lobiferus , P. pulitzerfinali , and P. spongicola ( Holthuis, 1952; Bruce, 1970, 1975, 1976, 1978, 2010, 2011; Fransen, 2006; Park & De Grave, 2021).
Of the remaining species, the new species can be easily differentiated from P. solitus by the number of dorsal rostral teeth (three vs. 11 in P. alius , see Bruce & Coombes, 1995); from P. aurantiacus by the same character (five teeth, see Bruce, 2014d); from P. mortenseni by the dactylus of the second chela with a biunguiculate tip (vs. simple in P. mortenseni ) as well as the presence of a poorly developed dorsal anteromedian lobe on the first tergite (absent in P. mortenseni , see Bruce, 1994); from P. hebedactylus by the position of the dorsal cuspidate setae on the telson (both pairs at about 0.2 of telson length in P. hebedactylus vs. proximal pair at about 0.1 and distal pair at 0.5 in new species) as well as the well-developed, dorsal anteromedian lobe on the first tergite (poorly developed in the new species, see Bruce, 2012); from P. matherae by the size and position of the antennal tooth in that species, entire cutting edge of the minor second chela (vs. serrated in P. matherae ), as well as the differential shape of the dactylus of the third pereiopod (see Bruce, 2005a); from P. pachyspinosus, P. rastrifer , and P. tchesunovi by the distally denticulate chelae of the first pereiopod (vs. simple in P. alius , see Bruce, 1980; Ďuriš, 1990; Marin, 2007); whilst the smooth, lateral margin of the uropodal exopod allows the new species to be easily distinguished from P. nufu , which has serrated margins ( Ďuriš et al., 2009).
Of the 19 species currently known from the Indo-Pacific region, which have the dactylus of the third pereiopod armed with a distal accessory tooth, the corpus of which has ventral denticles but with the unguis being non-dentate; the new species thus appears morphologically similar to only four species, which may not necessarily form a phylogenetic clade, viz. P. apomonosi , P. bidentatus , P. leptodactylus , and P. tuamotae . These species share the following characteristics: 1) multidentate rostrum with more than six teeth; 2) the presence of a supraorbital tooth or tubercle; 3) the relative position of both pairs of dorsal cuspidate setae on the telson (near proximal margin and mid-length); 4) the non-denticulate cutting edges of the major and minor second chelae; 5) the presence of tubercles along the ventral border of the merus of the second pereiopods; and 6) the presence of spiniform setae along the ventral margin of the third pereiopods.
The recently described P. apomonosi from Korea ( Park & De Grave, 2021) shares two further characters with the new species: the presence of a pointed process on the inferior orbital margin, and the presence of the dorsal anteromedian lobe on the first tergite. However, P. apomonosi is readily distinguished from the new species by the simple tip of the dactylus of the minor second chela (vs. bidentate in P. alius ), as well as further differences, such as proportions of the eye (about 1.3 times as long as maximal diameter in P. apomonosi vs. about 1.6 times in P. alius ), and the absence of a small acute median process of distal margin of the telson (present in P. alius ).
Periclimenaeus bidentatus is currently known from Australia (Queensland, Northern Territory, Western Australia), East Africa, Papua New Guinea, and New Caledonia ( Bruce, 2013). Based on the unillustrated type description ( Bruce, 1970) as well as the illustrations in Bruce (1991b), this species shares the presence of the bidentate tip on the dactylus of the minor second chela (see fig. 18f in Bruce, 1991b). However, P. bidentatus clearly differs from P. alius as follows: 1) shape of the inferior orbital margin (feebly developed and blunt in P. bidentatus vs. pointed process in new species); 2) length of distolateral of scaphocerite (far exceeding the distal lamella in P. bidentatus vs. reaching to end of distal lamella in P. alius ); 3) the length of carpus of the first pereiopod (about 1.5 times of chela in P. bidentatus vs. subequal to chela in P. alius ); 4) the ornamentation of the fourth and fifth ambulatory propodi (absent in P. bidentatus vs. present in P. alius ); and 5) the shape of the distal margin of the telson (median process absent in P. bidentatus vs. present, but minute, in P. alius ).
Periclimenaeus leptodactylus has only been reported from the Ryukyu Islands and East Africa ( Bruce, 2013). Based on the illustrated type description ( Fujino & Miyake, 1968), this species is readily separated from the new species as follows: 1) the shape of the inferior orbital margin (slightly produced in P. leptodactylus vs. pointed in P. alius ); 2) the shape of the outer margin of the antennal basicerite (unarmed in P. leptodactylus vs. with a pointed process in P. alius ); 3) the length of the antennal carpocerite (long, reaching to end of the distal article of the antennule peduncle vs. short, reaching to proximal article in P. alius ); 4) tip of the dactylus of the minor second chela simple (vs. bidentate in P. alius ); and 5) the dactyl of the first pereiopod with pectinations in about the distal half (vs. entire in P. alius ). A minute process on the distal margin of the telson could have been overlooked in the type description of P. leptodactylus , but if truly absent, could serve as a further diagnostic difference.
Periclimenaeus tuamotae is currently known from the Tuamotu Archipelago ( French Polynesia), East Africa, Sabah, and Australia (Northern Territory, Queensland) ( Bruce, 2013), although the only description (lacking illustrations) is by Bruce (1969). Based on the type description, that species differs from P. alius as follows: 1) smaller number of dorsal rostral teeth (six vs. 11 in P. alius ); 2) longer eye (extending beyond the tip of the rostrum vs. extending to about 0.8 of rostrum length in P. alius ); 3) the shape of the inferior orbital margin (obsolescent vs. pointed in P. alius ); 4) the distolateral tooth of the scaphocerite overreaching the distal margin of the lamella (vs. reaching to distal margin in P. alius ); 5) the unarmed shape of the outer margin of the antennal basicerite (vs. pointed process in P. alius ); 6) length of the carpus of the first pereiopod (about 1.5 times of chela in P. tuamotae vs. subequal to chela in P. alius ); and 7) the simple tip of the dactylus of the minor second chela (vs. bidentate tip in P. alius ). A minute process on the distal margin of the telson could have been overlooked in the type description of P. tuamotae , but if truly absent, could serve as a further diagnostic difference.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |