
Paranourosorex intermedius, Zazhigin & Voyta, 2022
|
publication ID |
https://doi.org/ 10.26879/1209 |
|
publication LSID |
lsid:zoobank.org:pub:1726FDAE-2EE5-4145-A124-6D24287C0514 |
|
DOI |
https://doi.org/10.5281/zenodo.11105202 |
|
persistent identifier |
https://treatment.plazi.org/id/71755174-FFA7-0336-CE0D-FD79CA83FD83 |
|
treatment provided by |
Felipe |
|
scientific name |
Paranourosorex intermedius |
| status |
sp. nov. |
Paranourosorex intermedius sp. nov.
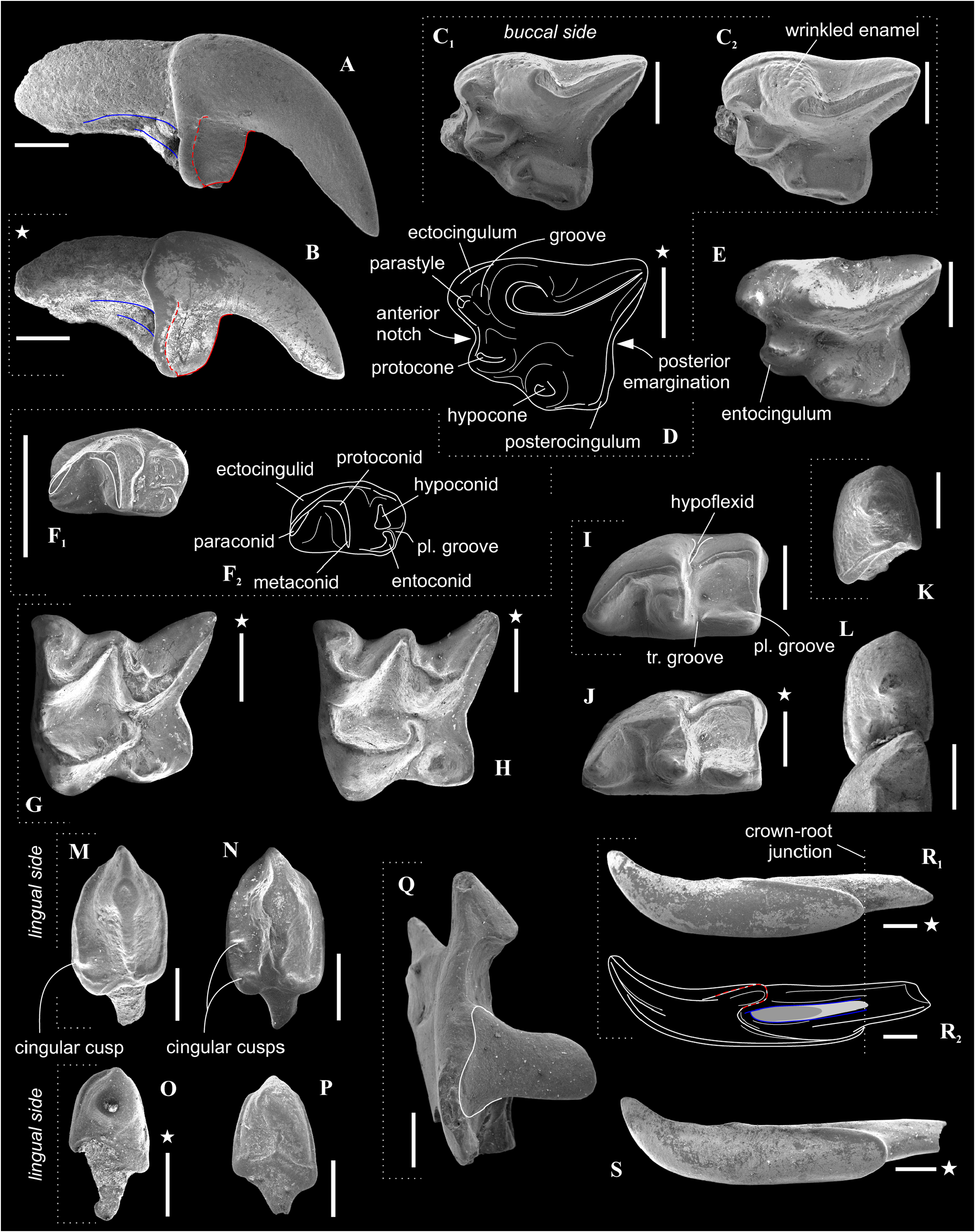
Figures 6E View FIGURE 6 , 7B, C–D, F–G, I, K, M, O, Q–R View FIGURE 7
zoobank.org/ FA6D006C-2C2A-4860-BE5A-BBAD043A2D49
1996 Paranourosorex sp. 2 Storch and Zazhigin, p. 264, figs. 3c, 4l–o.
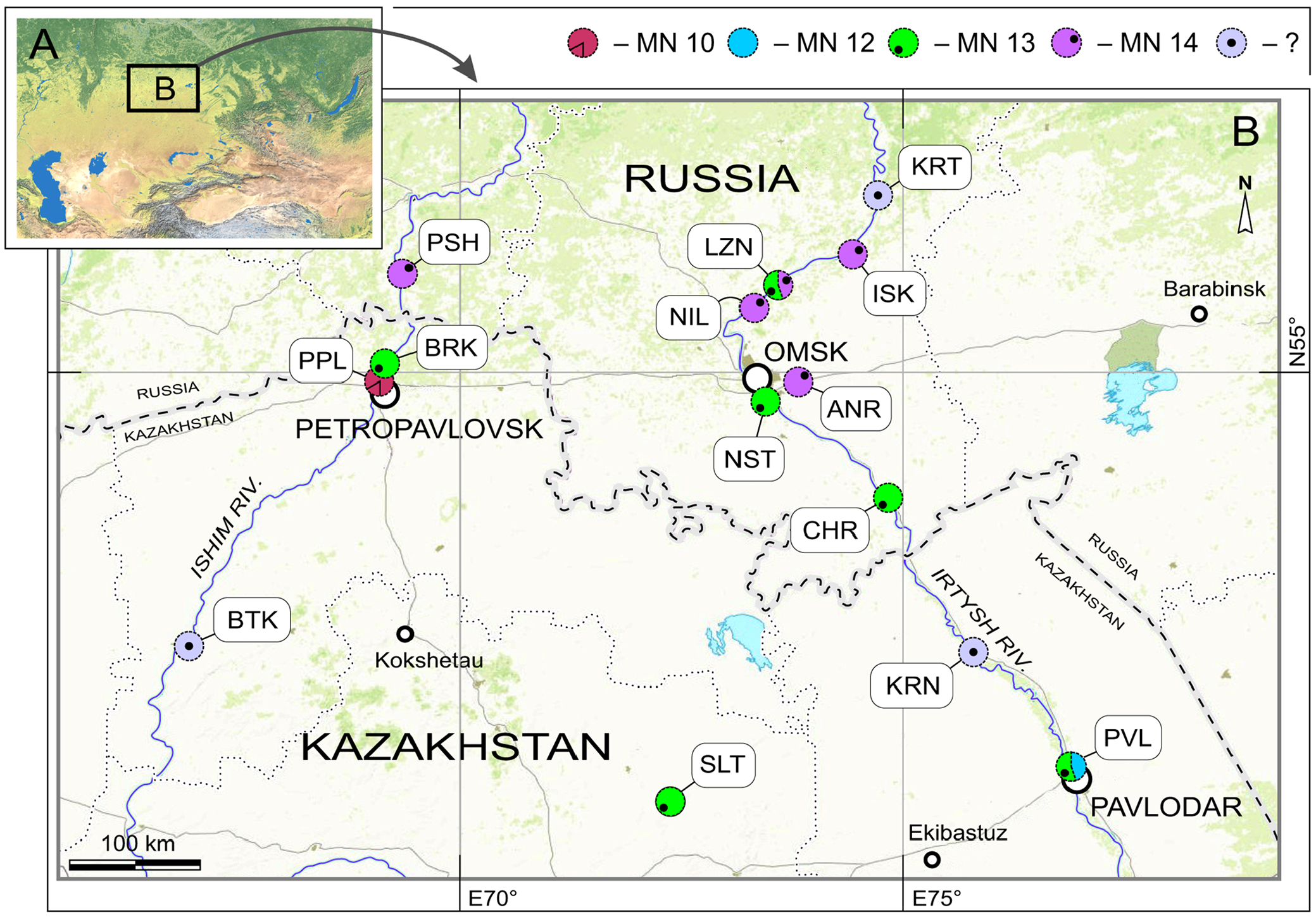
Type locality. Novaya Stanitsa GoogleMaps 1A, right slope of the Irtysh River Valley GoogleMaps near Novaya Stanitsa village (ca. N54°50' E73°24'), Omskaya Oblast’, Russia ( Figure 1 View FIGURE 1 : NST).
Type horizon. Stratotypic locality of Novaya Stanitsa Formation (nst), late Miocene (Turolian, MN 13). Material was collected by VZ during fieldworks of 1980s, 2000 and 2001.
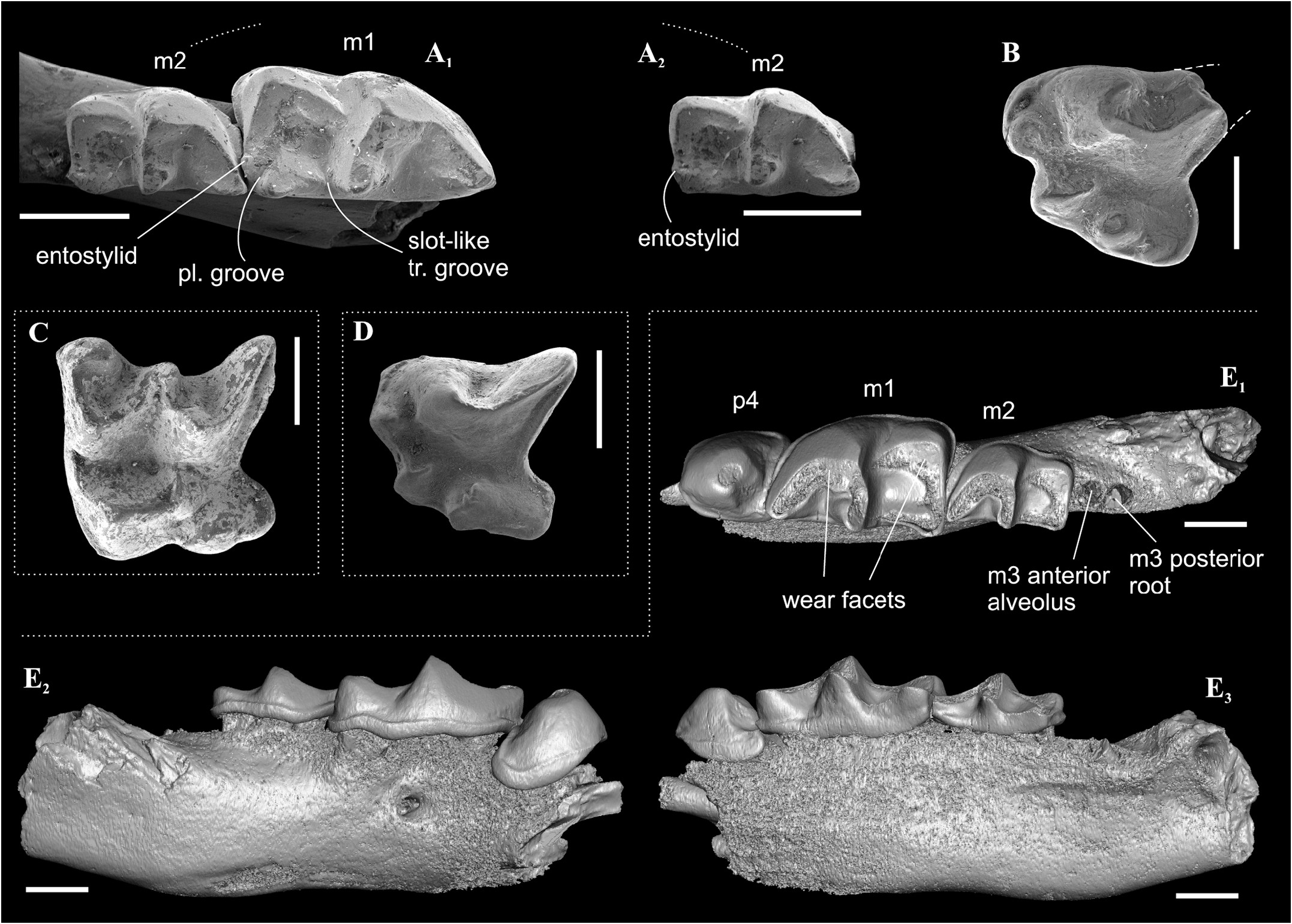
Type material. Holotype: GIN 948/1051 — right dentary fragment with moderately worn p4 and heavily worn m1–m3. Paratypes: (n =12): GIN 948/ 1052 — left dentary fragment with i1, m1 and alveoli of a1–p4 (i1 heavily and m1 moderately worn) ; GIN 948/1056 — isolated right m1 (slightly worn); GIN 948/1063 — isolated left m2 (slightly worn); GIN 948/1064 — isolated right m3 (slightly worn) ; GIN 948/1065 — isolated left P4 (slightly worn); GIN 948/1069 — isolated right M1 (metacone damaged; moderately worn); GIN 948/ 1070 — isolated right M1 (paracone and metacone damaged; moderately worn) ; GIN 948/1073 — isolated left I1 (slightly worn) ; GIN 948/1075 — isolated right i1 (tip of root is broken off; moderately worn); GIN 948/1078 — isolated left p4 (without roots; unworn) ; GIN 948/1081 — isolated left a1 (slightly damaged and worn) ; GIN 948/1089 — isolated left A1 (slightly worn).
Location of types. Holotype and paratypes in the collection of GIN, Moscow ( Russia).
Etymology. From the Latin intermedius , intermediate (between smallest P. seletiensis and largest P. gigas ).
Material. Type material and 27 remains from type locality (NST/1A), 60 remains from other 8 localities: NST/1B (n = 1), BRK/1A (n = 26), BRK/1B (n = 18), LZN/2B (n = 8), PVL/2 (n = 1), ISK/1A (n = 1), ISK/2A (n = 3), KRT (n = 1) (see Appendix 2: 3).
Measurements. See Appendix 4.
Diagnosis. Large-sized Paranourosorex species. Lower teeth, mostly p4 and m1, are inflated. Upper antemolar row consists of three teeth with vestigial A3. P4 has trapeziform shape with a deep anterior notch and very shallow the posterior emargination. M1 has subquadrate shape with the expressed Wshaped buccal crests, moderately expressed short metaloph without the metacone base contact. p4 is moderately exaenodont, bulbous-like in shape with the spot-shaped wear facet; the posterolingual basin of p4 is very weak. The entocristid of m1 is separated from the metaconid base by a distinctly marked transverse groove; the hypolohid is low; the small entostylid is weakly separated from the entoconid base.
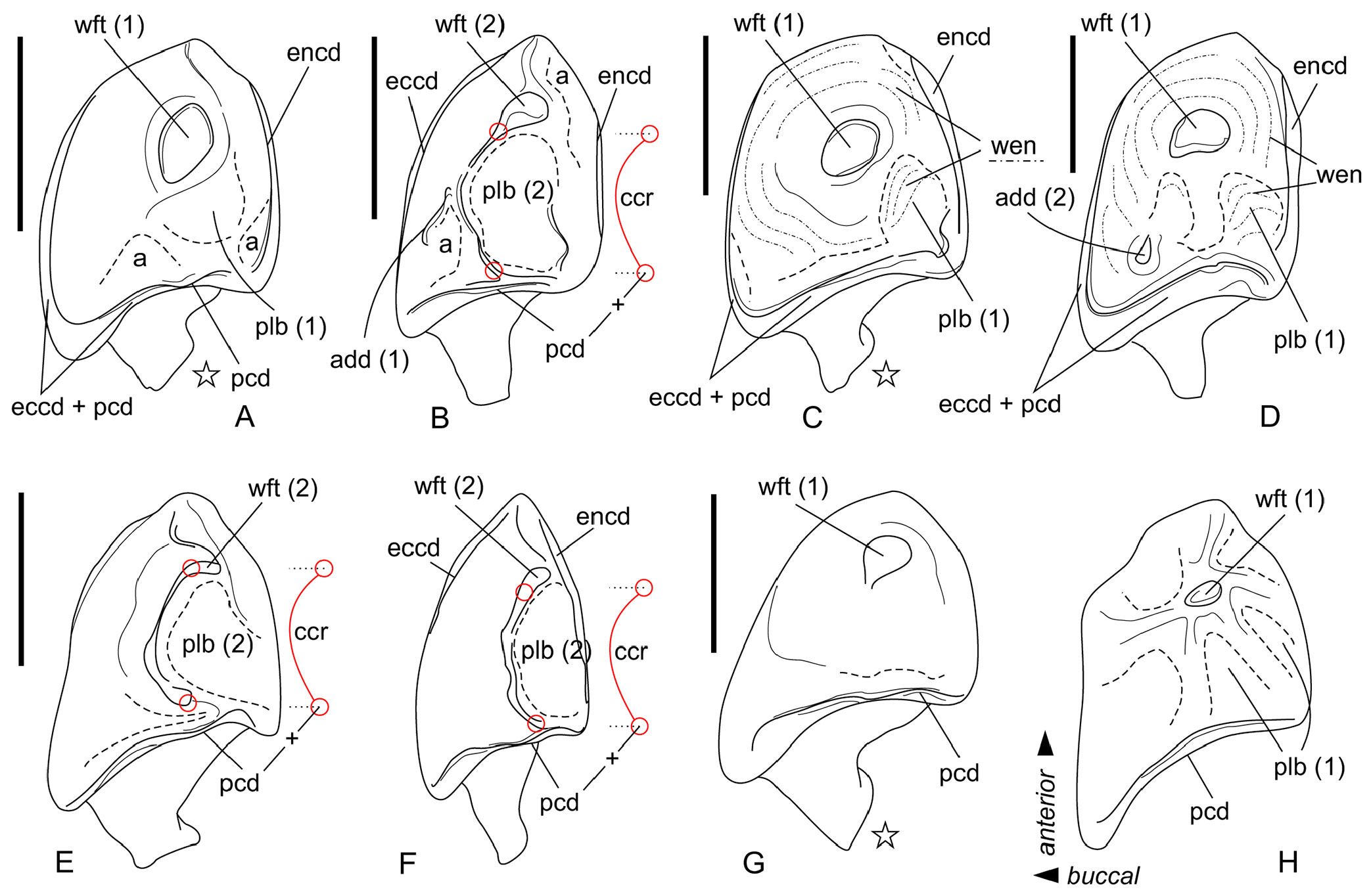
Description. I1 crown proportional in size to the root; the talon has an oblique tilted cutting edge without any notch in the base; the ectocingulum is moderately developed and rises up to the middle of the crown ( Figure 7B View FIGURE 7 ). The upper antemolars are dramatically decreased in size from the largest A1 to middle-sized A2 (less than 1/2 of A1) and vestigial A3. The addition of cingular cusps (usually one cusp) on the lingual part of the crown is developed on A1; the entocingulum of A2 is smooth. A1 and A2 have developed both anterior and central crests; A1 displays a wrinkled enamel ( Figure 7M View FIGURE 7 ). P4 has trapeziform crown, developed anterior notch and the shallow posterior emargination. The parastyle is separated from the paracone base by a shallow groove. P4 has a short ectocingulum, well-developed postcingulum and wrinkled enamel on the paracone buccal side ( Figure 7C View FIGURE 7 ); the entocingulum is absent. M1 has subquadrate occlusal shape with the expressed W-shaped line of the buccal crests. The metaconule is absent; the metaloph is short and separated from the base of the metacone ( Figure 7G View FIGURE 7 ). The posterior emargination of M1 is shallow. M2 has well-developed cones; the parastyle is longest, the mesostyle is buccally prominent, the metastyle undeveloped; the hypocone round in the base with a spot-shaped wear facet. M3 unknown. i1 has a smooth cutting edge and distinctly upturned tip. The ectocingulid of i1 is undeveloped. The medial groove of the root is short and did not overreach the crown-root junction. a1 has a bulbous-like crown and massive root; the crown with well-developed ecto- and entocingulids, spot-shaped wear facet and short anterior crest. The central crests of a1 is undeveloped. The inflated p4 bears the spot-shaped wear facet, extremely weak developed the crown structures and distinctly wrinkled enamel ( Figures 4C View FIGURE 4 , 7K View FIGURE 7 ). m1 is relatively short and wide and so seems slightly inflated. The enamel surface of the trigonid buccal side is slightly wrinkled. The protoconid and metaconid of m1 are brought closer, the hypoconid and entoconid are widely separated. The hypoflexid of m1 is a deep almost vertical groove that does slightly not reach the ectocingulid. The hypolophid is worn in different degrees in all examined specimens, but despite this, we can see two morphotypes: the hypolophid is weakly separated from the entoconid, the entostylid is absent and the posterolingual groove very weak (Morphotype I, GIN 948/1056, NST/1A) ( Figure 7I View FIGURE 7 ); the hypolophid is clearly separated from the entoconid, the small entostylid and the shallow and short posterolingual groove are present (Morphotype II, GIN 1115/1126, BRK/1B). The entoconid is high and relatively narrow; the entocristid is separated from the metaconid base by a narrow transverse groove. The ectocingulid is well-distinguished along the base of the tooth; entocingulid is absent. m2 notably less than m1; the crown bears the high trigonid and the low talonid. The entostylid is absent; the hypolophid contacts the entoconid without the groove. The entocristid is separated the base of the metaconid by a narrow slot. The ectocingulid m2 is very well-developed along the tooth base with the plate-like extension in the first third (paraconid level). The small m3 has the well-developed trigonid and the reduced talonid without a talonid basin; the talonid consists of the small hypoconid and entoconid that separated by the shallow posterolingual groove ( Figure 7F View FIGURE 7 ). The ectocingulid of m3 bears the plate-like extension in the first third that is similar to m2.
A single left premaxilla-maxilla fragment with I1 and A2 from PSH /1B (GIN 1118/1027) was examined. I1 is moderately worn; double-rooted A2 is slightly worn. Two alveoli of A1 indicate the presence of the small separated anterior root of A1. Thus, A1 and A2 are both double-rooted.
The mandibular body is long and relatively low without a visible inward curve of the lower outline. The mental foramen is positioned under the protoconid of m1 without any grooves in a wide and shallow depression. The mandibular ramus is massive with a small internal fossa for the temporal muscle; its maximal length is lesser than the condylar height (HC). The coronoid process includes a well-developed long coronoid spicule. The single mandibular foramen is large. The lower articular facet of the condylar process shifts far anteriorly; an interarticular area is relatively narrow ( Figure 7Q View FIGURE 7 ). An angular process is unknown.
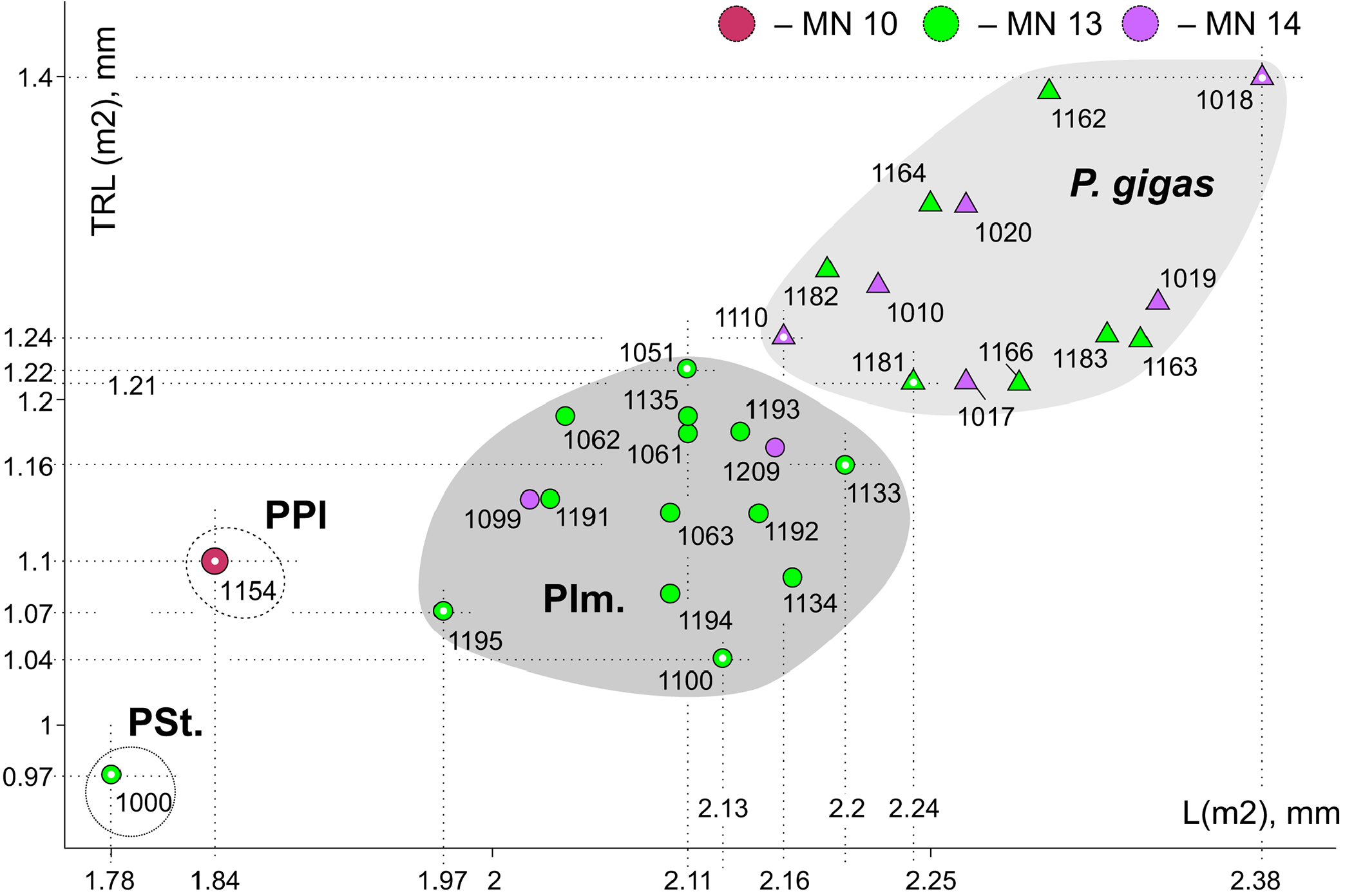
Comparisons. Paranourosorex intermedius sp. nov. differs from P. seletiensis in a larger size ( Figure 5 View FIGURE 5 ; Appendix 4) and some dental characters (the first species has longer hypoconal flange of P4 and more inflated m1 and m2 than has the second species; Figure 6B View FIGURE 6 cf. Figure 7D View FIGURE 7 ). Paranourosorex intermedius sp. nov. differs from P. inexspectatus in a slightly larger size; in the characters of P4 such as a more developed anterior notch and a more shallow posterior emargination, absent the entocingulum; in a more developed metastyle of M1 (the metastyle of P. inexspectatus is relatively shorter and narrower); in a more lingual position of the hypocone and a slightly deeper posterior emargination of M1 (for P. inexspectatus see Storch, 1995: plate 4). Paranourosorex intermedius sp. nov. differs from P. gigas in a slightly smaller size; in a more shallow anterior notch and posterior emargination of P4 ( Figure 7 View FIGURE 7 : C cf. E); in a more developed metastyle of M1 (the metastyle of P. gigas is relatively narrower); in a more lingual position of the hypocone, a slightly longer hypoconal flange and a slightly deeper posterior emargination of M1; in absence of the entocingulum of M1 ( Figure 7 View FIGURE 7 : G cf. H); in strongly exaenodont buccal half of a1 ( Figure 7 View FIGURE 7 : O cf. P); in presence of a weak posterolingual groove of m1 ( P. gigas has not the groove, the hypolophid contacts the entoconid; Morphotype I).
Remarks. The fossils of Paranourosorex intermedius sp. nov. correspond to ‘ Paranourosorex sp. 2 ’ in Storch and Zazhigin (1996: 264). That description comprises several discordant features that require clarification. Authors stated: “its [p4] apex is weakly two-cusped.” We re-evaluated the apex of p4 as a distinctly single cusp with a spot-shaped wear facet ( Figure 7K View FIGURE 7 ). Authors also stated: “the hypolophid on M/1 is separated from the entoconid by a groove (even in worn specimens).” We found two morphotypes of hypolophid/entoconid of m1 state when the hypolophid is weakly separated from the entoconid with a very weak groove, either the hypolophid is clearly separated from the entoconid with the small entostylid and the shallow and short posterolingual groove.
Much material of Paranourosorex intermedius sp. nov. originates from NST/1A (n = 39), BRK/1A (n = 26) and BRK/1B (n = 18). The species is more poorly represented in the other nine sites (Appendix 2). Paranourosorex intermedius sp. nov. probably coexisted with larger shrew P. gigas in CHR/1A (Turolian, MN 13, Rytov Formation), ISK / 1A (Ruscinian, MN 14, Isakov Formation), PSH /1A and PSH /1B (Ruscinian, MN 14, Peshnev Formation) localities.
Stratigraphic and geographic range. At present, the distribution includes the type locality and 11 localities from Kazakhstan and Russia (Appendix 2) between the late Miocene Novaya Stanitsa Formation (Turolian, MN 13) and the early Pliocene Peshnev Formation (Ruscinian, MN 14).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Soricomorpha |
|
Family |
|
|
SubFamily |
Soricinae |
|
Tribe |
Anourosoricini |
|
Genus |