
Paranourosorex gigas Rzebik-Kowalska, 1975
|
publication ID |
https://doi.org/ 10.26879/1209 |
|
publication LSID |
lsid:zoobank.org:pub:1726FDAE-2EE5-4145-A124-6D24287C0514 |
|
DOI |
https://doi.org/10.5281/zenodo.11105204 |
|
persistent identifier |
https://treatment.plazi.org/id/71755174-FFA4-0336-CFC5-FDFBCC92FCC3 |
|
treatment provided by |
Felipe |
|
scientific name |
Paranourosorex gigas Rzebik-Kowalska, 1975 |
| status |
|
Paranourosorex gigas Rzebik-Kowalska, 1975
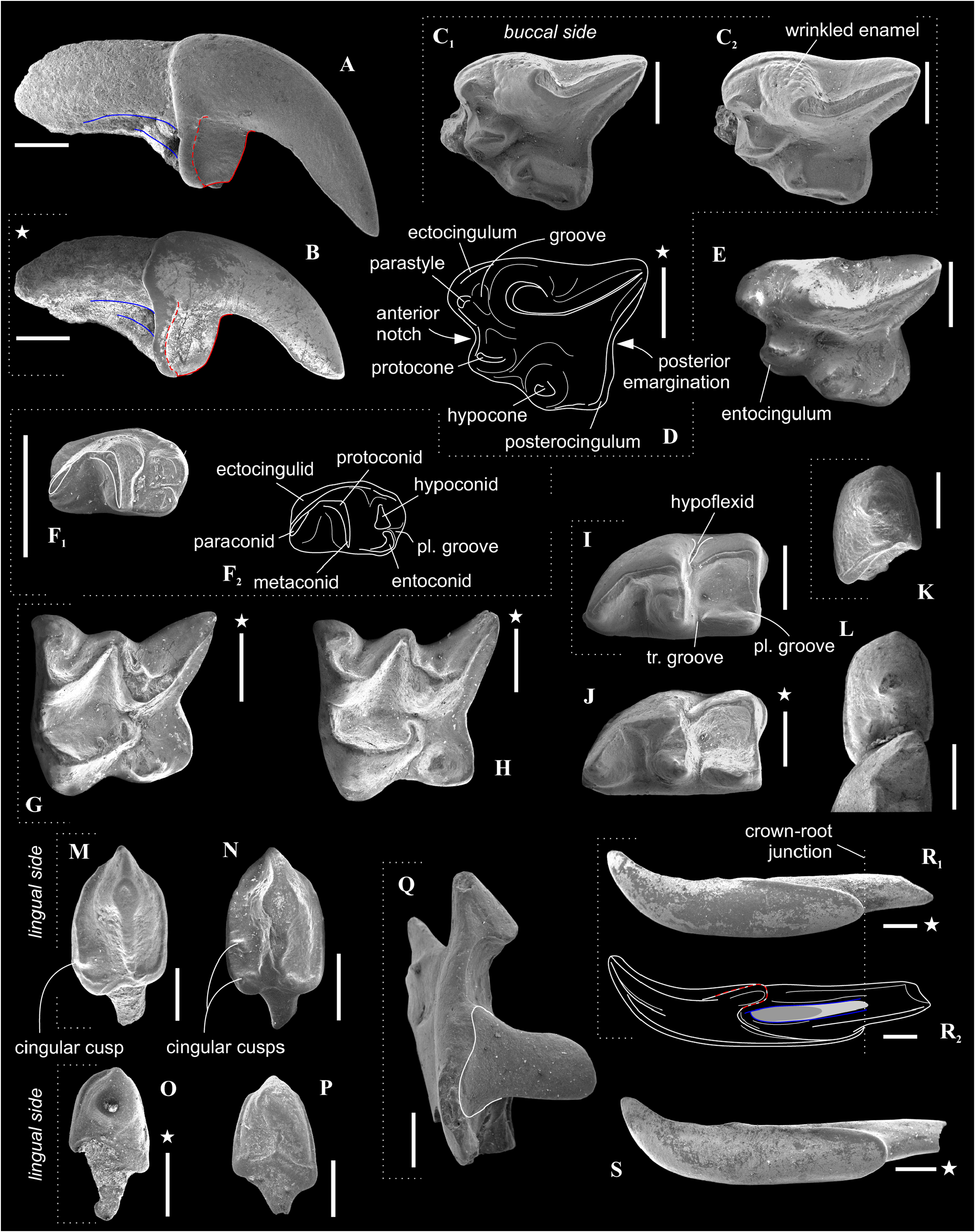
Figure 6A, E, H, J, L, N, P, S View FIGURE 6
1978 Crocidura sp. Agadjanian and Kowalski, p. 33.
1988 Paranourosorex sp. Topachevsky et al., p. 20.
2009 Soricidae gen. Agadjanian, p. 102.
Material. The analysed material of P. gigas comprises remains mainly from CHR/1A (n = 20) and PSH /1B (n = 33) localities . Single remains originate from ISK /1A (n = 1) , NIL (n = 2) , PSH /1A (n = 1), ANR/1A (n = 2) , ANR/2A (n = 1) , LZN/1A (n = 3) , BRK/1C (n = 1) and KRN (n = 2) localities (see Appendix 2: 4).
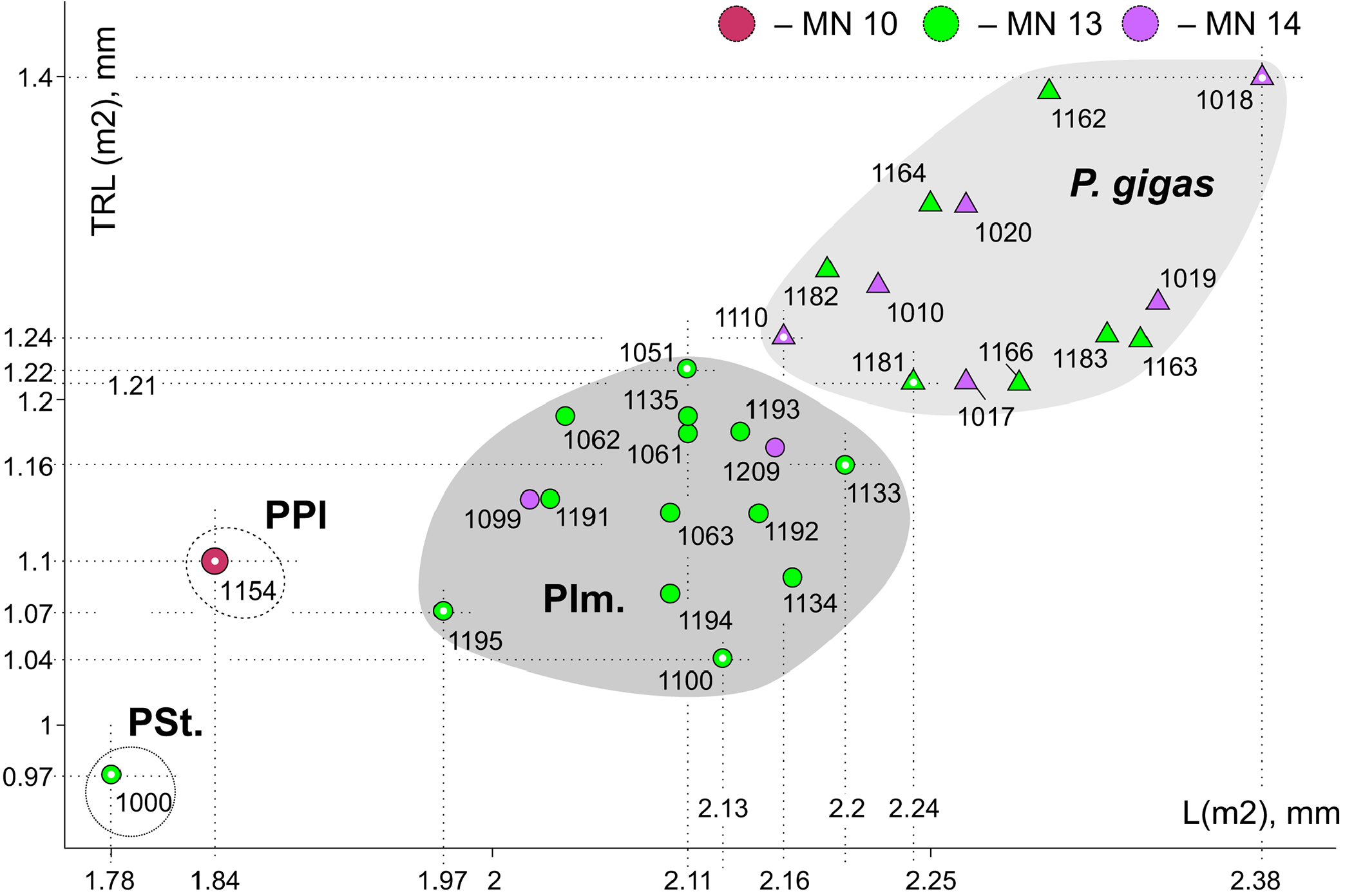
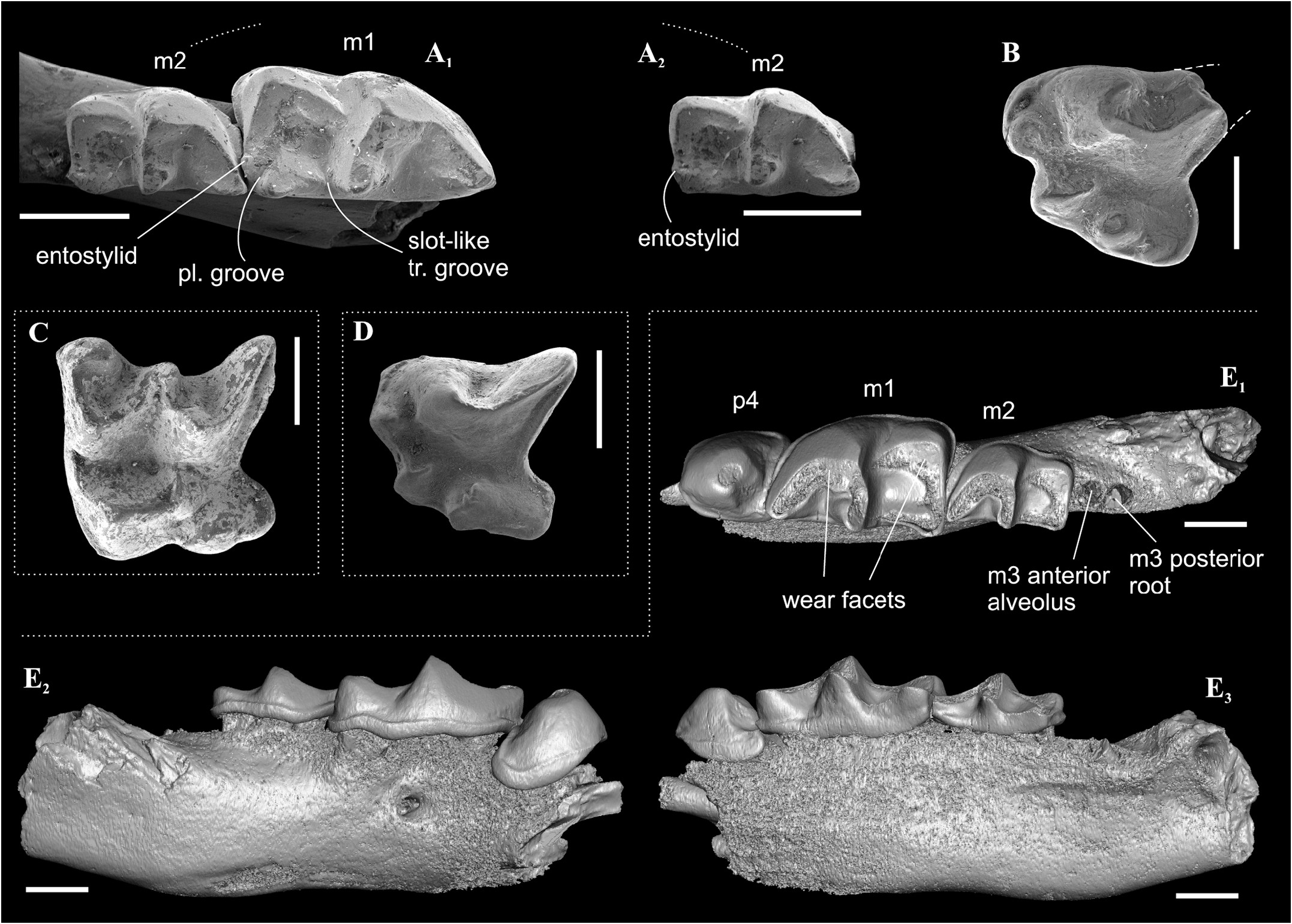
Description. Largest Paranourosorex species. In general, the dental characters are typical for the genus but some teeth display specific characters that distinguish this species such as the most undulated and sculptured P4 outline between the parastyle and hypocone with the well-developed anterior notch, the short portion of the entocingulum under the protocone, the moderately developed notch before the hypocone and the protocone lobe; the relatively deep posterior emargination of P4 ( Figure 7E View FIGURE 7 ); the moderately developed metastyle of M1; the massive cone-like hypocone of M1, relatively short the hypoconal flange and the shallow posterior emargination of M1 ( Figure 7H View FIGURE 7 ); M1 has a weak entocingulum under the protocone; the moderately exaenodont a1 has a wide postcingulid ( Figure 7P View FIGURE 7 ); the hypolophid of m1 contacts the entoconid base without a groove; a tip of the entoconid of m1 is slightly shifted anteriorly; the entocristid of m1 is separated from the metaconid base through the moderately developed transverse groove. m3 is unknown.
Measurements. See Appendix 4.
Remarks. P. gigas is the biggest species of the tribe, especially in comparison to P. seletiensis ( Figure 5 View FIGURE 5 ). To differentiate P. gigas from Paranourosorex intermedius sp. nov. see above. VZ has seen materials from Obukhovka 1 referred to as ‘ Paranourosorex sp. ’ ( Topachevsky et al., 1988); from Antipovka and Chugunovka referred to as ‘ Crocidura sp. ’ by Agadjanian and Kowalski (1978) and ‘ Soricidae gen.’ by Agadjanian (2009) and determined all findings as P. gigas .
Much material of P. gigas originates from CHR/1A (n = 20) and PSH /1B (n = 33).
Stratigraphic and geographic range. At present, the distribution includes the type locality Podlesice (early Pliocene , Ruscinian, MN 14, Poland), Slovakian locality (early Pliocene , Ruscinian, MN 15) ( Rzebik-Kowalska , 1975, 1998), three Eastern European Plain localities such as Obukhovka 1 ( Topachevsky et al., 1988), Antipovka and Chugunovka ( Agadjanian and Kowalski , 1978; Agadjanian , 2009; early Pliocene , Ruscinian, MN 14) and also 10 localities from Kazakhstan and Russia ( Appendix 2). The stratigraphic range in Asia covers an interval between the late Miocene Rytov Formation (Turolian, MN 13) and early Pliocene Peshnev Formation (Ruscinian, MN 14).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |