
Paramysis (Longidentia) adriatica, Wittmann, Karl J., Ariani, Antonio P. & Daneliya, Mikhail, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4142.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:FA423164-276C-44B0-A417-8E97AC3DF0AA |
|
DOI |
https://doi.org/10.5281/zenodo.6088653 |
|
persistent identifier |
https://treatment.plazi.org/id/03B9BA4E-181A-FFC5-CAF6-FEEDFEF0462B |
|
treatment provided by |
Plazi |
|
scientific name |
Paramysis (Longidentia) adriatica |
| status |
sp. nov. |
Paramysis (Longidentia) adriatica sp. nov.
Figs 7 View FIGURE 7 A, B, 8–10, 11A
Mysis oculata relicta: Stammer 1932 View in CoL (partim: station Tb9 ").
Paramysis helleri: Holmquist 1955 View in CoL ; Ariani et al. 1993: Tab. I (partim: Split); Ariani 2004: fig. 3B.
Type material. Holotype male with 9.1 mm body length ( NHMW reg. no. 25694), 21 paratypes (16 F ad. 9.3–11.7 mm, 4 M ad. 7.9–8.9 mm, 1 F subad., NHMW 25695 View Materials , MZH HLA. 112100, MZUN, SMNH Type 8783, MFSNU) (co-occurring with eight Diamysis fluviatilis ), NW-Adriatic coast, Adige River , 9 km from its mouth into the sea, 45.1160N 012.2684E, altitude 0 m, 0.4–0.8 m depth, among flooded bank vegetation after several days of rain, roots and branches of Salix , v = 0.05 m /s, S = 0.1, 305 µS/cm, 10.6°C, pH 5.48, 8.27 mg O2/l, 8°d, 37 NTU GoogleMaps , 14 April 2012, hand net, leg. Ariani & Wittmann.
Additional material examined (stations listed in clockwise order along Adriatic coasts). 4 F ad. 9.8–11.5 mm, 2 imm., NW-Adriatic coast, Po di Goro = mouth branch of Po River, at Mesola, 44.9251N 012.2310E, 27 km to the sea, altitude 2 m, 0.2–0.7 m depth, flooded bank vegetation, Salix , concrete, v = 0.03–0.06 m /s, S = 0.1, 442 µS/cm, 13.3°C, pH 6.43, 7.14 mg O2/l, 8°d, 81 NTU, hand net, 14 Apr. 2012, leg. Ariani & Wittmann, NHMW 25697 View Materials GoogleMaps ; 1 F ad. 9.3 mm, 1 F imm. 8.7 mm, N-Adriatic coast, near Panzano , Canale Brancolo, 45.7804N 013.5129E, 4 km to the sea, altitude 0 m, benthic, 29 Sept. 1986, leg. F. Stoch & B. Zanolin, MFSNU; dissected parts of 1 M ad. 7.5 mm, 1 F ad. 8.5 mm with 2 postnauplioid larvae, on a total of 2 slides labelled " Paramysis helleri . Timavo. Det. Ch. Holmquist. prep. 64, 65", SMNH reg. nos GoogleMaps . 140106, 140107, according to Stammer (1932) and Holmquist (1955: Tab. 1) this material was collected in 1928/29 by Hans Jürgen Stammer in the former right mouth branch of the Timavo , 45.7839N 013.5733E, Gulf of Trieste, NE-Adriatic; 1 F ad. 8.8 mm, 1 F imm. 6.6 mm, E-Adriatic coast, small freshwater creek at Split, roughly 43.515N 016.541E, 0–1 m depth, macrophytes, hand net, 26 Oct. 1976, leg. A. P. Ariani, NHMW 25698 View Materials GoogleMaps ; 6 M ad. 8.3–9.4 mm, 4 F ad. 9.4–10.9 mm, 2 F subad., 1 F imm., E-Adriatic coast, lake Deran (= Deransko jezero), oligotrophic karstic lake, 43.04N 017.75E, altitude 0 m, total of about 31 km along its small effluent and following this along the Neretva River to the Adriatic Sea, sample labelled ”in clean and cold stagnant water in areas with mud bottom, specimens caught over the fine muddy sediment without any cover”, 0.5– 1 m depth, S = 0.2, 13–15°C, 9 July 2009, leg. D. Lučić & P. Tutman, NHMW 25696 View Materials GoogleMaps .
Derivatio nominis. The species name is a Late Latin adjective with female ending, referring to the so far known exclusive occurrence of the new species in tributaries of the Adriatic Sea.
Type locality. NW-Adriatic drainage basin, in fresh-water of the lower reach of the Adige River, 45.12N 012.27E ( Fig. 12 View FIGURE 12 ). This is the second-largest tributary of the Adriatic Sea.
Occurrence ( Fig. 12 View FIGURE 12 ). In freshwater tributaries of the Adriatic Sea. So far known only from five running waters and one lake, in each case less than 5 m above sea level; maximum sea distance 31 km. This enumeration includes the former right mouth branch of the Timavo, now dry land. Whether Stammer (1932) found any Paramysis in other parts of the Timavo system cannot be judged currently because there is no Timavo material in the Stammer collection at the Department of Biology, University of Erlangen, Germany (Wolfgang Heimler, pers. comm.). The Swedish Museum keeps only those Timavo materials inspected by Holmquist (1955) and 60 years later by us. Inspection of Timavo springs and estuary by one of us (KJW) in Sept. 1987 yielded no Paramysis materials.
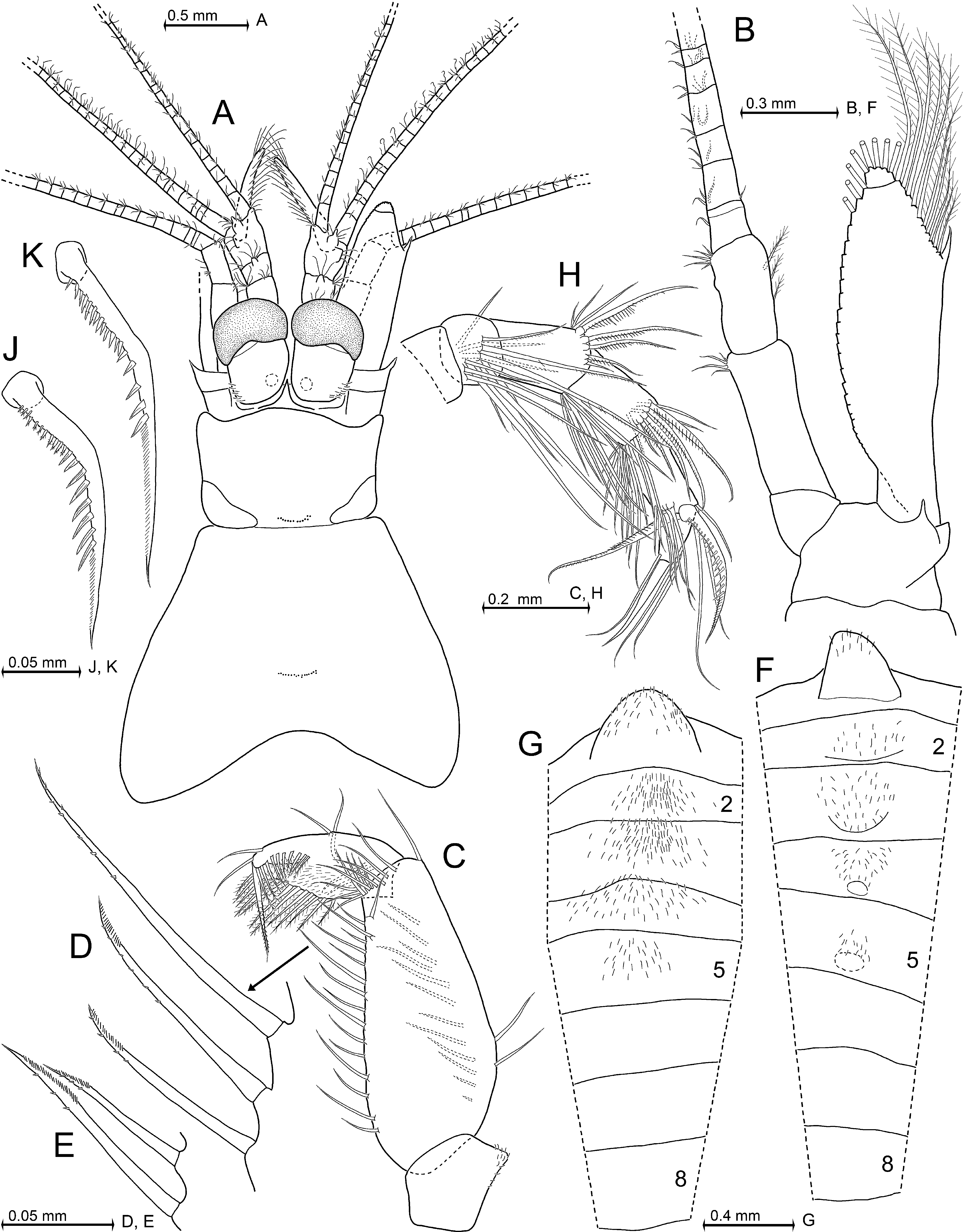
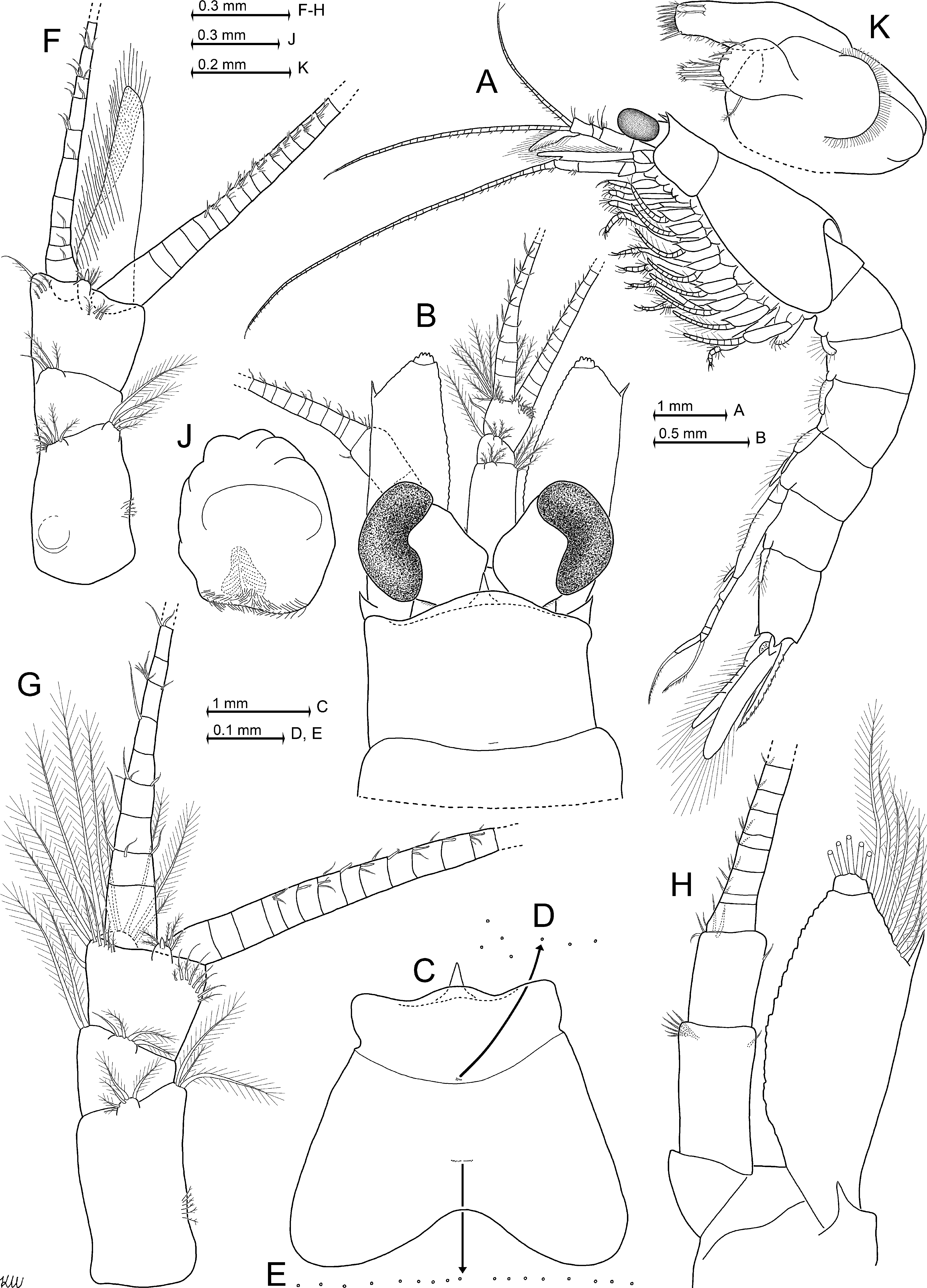
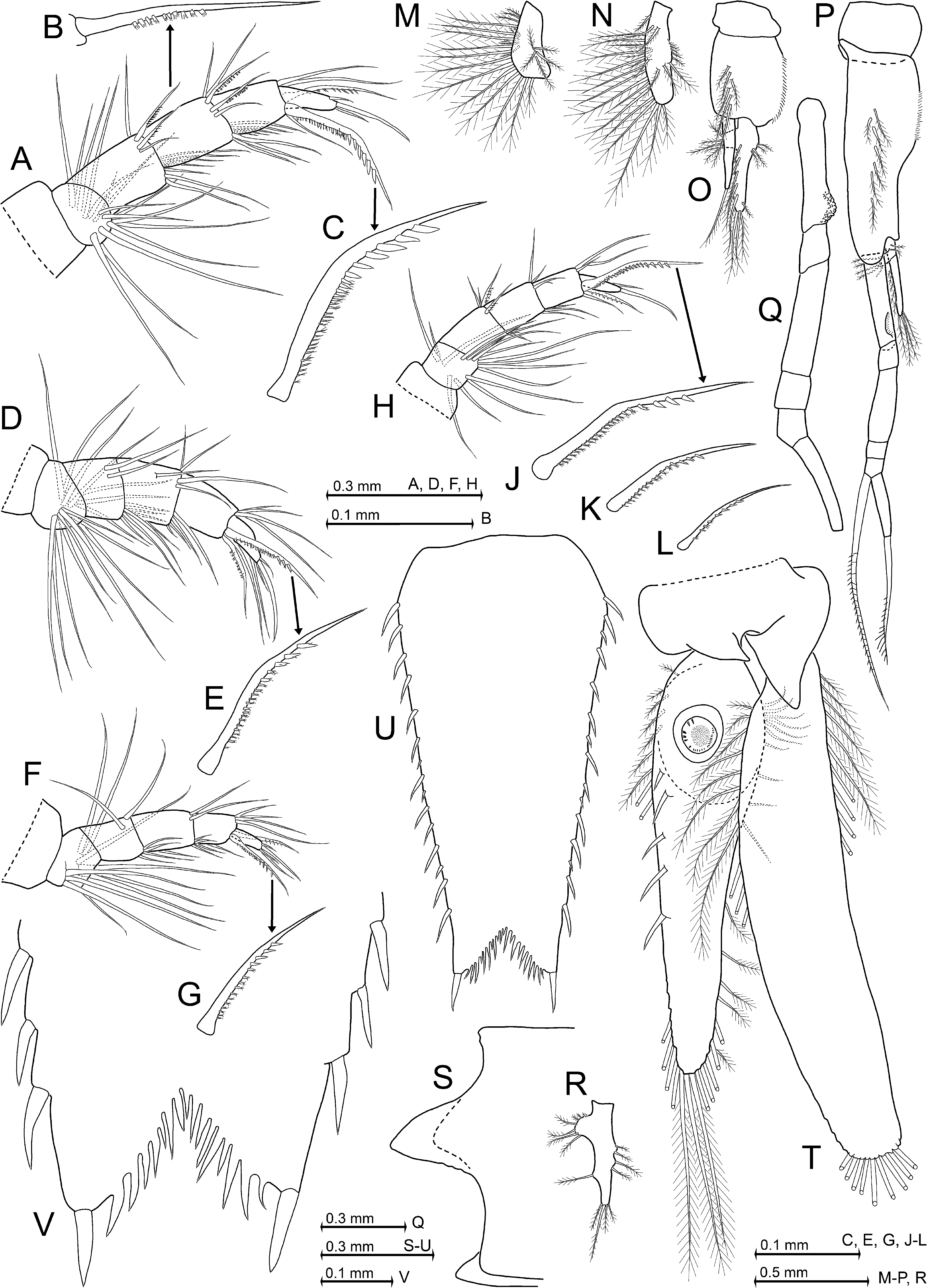
Definition. Antennal scale length is 2.5–2.6 times its maximum width; scale with smooth outer margin ending slightly before or at the terminal margin of antennular trunk or more often extending beyond this margin ( Fig. 8 View FIGURE 8 B), in any case ending at a non-articulated spiniform process; setose apex is 15–25% length of scale; apex bears a small apical segment ( Fig. 8 View FIGURE 8 H). Eyes with large, reniform cornea ( Fig. 8 View FIGURE 8 B); maximum diameter of cornea is 42–53% antennal scale length; ventral face of eyestalks with traverse stripe of hair-like scales. Wedge- to sickle-shaped, mostly upward-bent, subrostral process extends well beyond the short, evenly rounded rostrum ( Fig. 8 View FIGURE 8 B, C). This process appears subtriangular in dorsal view ( Fig. 8 View FIGURE 8 B). Carapace with medium-sized, subtriangular, basally rounded, mid-dorsal, posterior emargination ( Fig. 8 View FIGURE 8 C). Median segment of mandibular palp with normal (i.e. not microserrated) setae ( Fig. 9 View FIGURE 9 A). Thoracic sternites 2–5 of adult males with median, apically rounded processes of different size, terminally equipped with hairs (hair-like scales); sternites 6–8 only with smooth, less distinct humps ( Fig. 9 View FIGURE 9 D, E); no such processes in adult females. Thoracic endopods 3–8 with 5-segmented tarsus ( Figs 9 View FIGURE 9 G, H, 10A, D, F, H); carpus as typical for the genus; propodus 3-segmented; dactylus minute, cylindrical, with small setalike nail. Endopod 3 with inner paradactylar seta ( Fig. 10 View FIGURE 10 C, J) showing a heavy armature with 4–6 spinules (i.e. stiff secondary projections) along its median to subterminal portions; proximal spinules thinner, their (sub)terminal portions in part with secondary 'hairs'; outer paradactylar seta with much weaker armature ( Fig. 10 View FIGURE 10 A, H). Penis with row of 4–6 short, barbed setae in subterminal position on caudolateral blade ( Fig. 9 View FIGURE 9 H; same type of setae as in P. kosswigi , Fig. 5 View FIGURE 5 A). Pleopods of both sexes (of males in Fig. 10 View FIGURE 10 M–R) as described above for the genus; male pleopods 3 ( Fig. 10 View FIGURE 10 O) and 4 ( Fig. 10 View FIGURE 10 P) each with unsegmented endopod; male pleopod 4 with 6-segmented exopod ( Fig. 10 View FIGURE 10 P, Q). Endopod of uropods with only 3–6 spines along inner margin between statocyst and 35–45% endopod length from tip ( Fig. 10 View FIGURE 10 T). Statoliths composed of vaterite. Telson with weakly curved, allusively Sshaped lateral margins; margins with 13–15 spines, not counting the (not or) moderately larger apical spines ( Fig. 10 View FIGURE 10 U, V). As an unusual state among species of Paramysis , shorter spines may alternate with longer ones on lateral margins of telson ( Fig. 10 View FIGURE 10 U; this pattern distinct in 18 out of 30 specimens examined, indistinct or missing in the remaining ones; terminal spines not considered). Triangular terminal cleft is 9–12% telson length, cleft not or barely reaching the subterminal spines ( Fig. 10 View FIGURE 10 U, V); cleft with almost straight margins on each symmetrical half, cleft armed with a total of 14–28 laminae of in part unequal size, even the longest laminae shorter than the subterminal or terminal spines, respectively.
Description (types only). All features within the ranges indicated above for the genus. Appendage morphology and habitus of both sexes ( Figs 7 View FIGURE 7 A, B, 8A, B) very similar to those of P. a gi ge n s i s and P. hel leri . Carapace leaves 1.5–2 posterior thoracic somites dorsally exposed. Carapace with traverse row of 16–22 pores in cardial position (above the heart), arranged in two roughly symmetrical subgroups, and 7–10 pores in cervical position ( Fig. 8 View FIGURE 8 C–E). Eyes dorsoventrally flattened. The cornea is reniform in dorsal view ( Fig. 8 View FIGURE 8 B), ovoid in lateral view ( Fig. 8 View FIGURE 8 A). Male antennula with slender appendix masculina facing obliquely downwards in the vertical plane ( Fig. 8 View FIGURE 8 A) and forwards in the horizontal plane ( Fig. 8 View FIGURE 8 F). This appendix is 2.0–2.3 times the length of apical segment of antennular peduncle. Sympod of antenna produced into a large spiniform process at its outer margin ( Fig. 8 View FIGURE 8 H). Antennal scale ( Fig. 8 View FIGURE 8 B, H) is 1.1–1.5 times the length of antennular peduncle ( Fig. 8 View FIGURE 8 A, B, F, G). Large non-articulated spiniform process (thorn) at 75–90% scale length from basis. Small terminal segment of scale with five large plumose setae. Basal segment of mandibular palp without setae, but with dense field of scales ( Fig. 9 View FIGURE 9 A). Remaining two segments setose. Median segment with smooth setae in both sexes; with a minor exception in males only, insofar as the basal three setae on the outer margin bear up to four subbasal to median barbs each ( Fig. 9 View FIGURE 9 A). Terminal segment half as long as the median one. Exopod of maxilla roughly fan-shaped, with 12–16 mostly long, plumose ( Fig. 9 View FIGURE 9 C) setae; exopod not larger than distal segment of palpus (= endopod). This palpus with terminal segment laterally widened, about 1.7–1.8 times wider than long.
First thoracic sternite produced into an anteriorly directed, distally hairy, medial lobe in both sexes ( Fig. 9 View FIGURE 9 D). Unlike the sterna of males given in the definition above, the sternites 2–8 of females are medially smooth, without any humps or hairs. Basis of all thoracic exopods laterally expanded, blade-like; its distal outer corner with spinelike extension in thoracopods 1–7 ( Fig. 9 View FIGURE 9 D), whereas well-rounded in thoracopod 8 ( Fig. 9 View FIGURE 9 H). Flagellum of exopods 1 and 8 each with eight segments, whereas in exopods 2–7 normally with nine segments, not counting the large intersegmental joint between basis and flagellum which may be mistaken as a segment. In contrast, the flagellum of exopod 7 showed only eight segments in one out of ten females inspected in this respect. Thoracic endopods rather stout, moderately long. Total length increases from endopods 1 to endopod 3, and then decreases backward to endopod 8. Endopod 8, when stretched anteriorly, extends only up to basis of endopod 3. First thoracic epipod with groups of minute scales along its anterior margin and on intersegmental joint with its sympod ( Fig. 9 View FIGURE 9 D). This sympod with additional groups of such scales and two plumose setae near insertion of epipod. First thoracic endopod with non-setose endite from coxa, and with large, setose endites from basis, ischium, and merus ( Fig. 9 View FIGURE 9 D). Ischium shorter than merus in thoracic endopods 1 and 2, slightly longer in endopods 3 and 8, or distinctly longer in endopods 4–7. Paradactylar setae of endopods 3–8 weakly sickle-shaped, their armature with denticles is stronger in females ( Fig. 10 View FIGURE 10 C) compared to males ( Fig. 10 View FIGURE 10 J) and decreases successively in both sexes from endopods 3 to 8 (females Fig. 10 View FIGURE 10 C, E, G; males Fig. 10 View FIGURE 10 J–L). Penis as in Fig. 9 View FIGURE 9 H. Exopod of fourth male pleopod ( Fig. 10 View FIGURE 10 Q): basal segment with strong, cuticularized ridge on its rostral face; this ridge more rugged than the homologous one found on the second segment in P. kosswigi ( Fig. 5 View FIGURE 5 D); the second segment in P. a dr i at i c a ( Fig. 10 View FIGURE 10 Q) with less strongly oblique segmental borders compared with the homologous third segment in P. kosswigi ( Fig. 5 View FIGURE 5 D). Scutellum paracaudale subtriangular with rounded tip, margins sinusoid, lower margin inconspicuously undulate ( Fig. 10 View FIGURE 10 S). Endopod of uropods is 77–84% length of exopod; spine size along inner margin of endopod and space between these spines increase, with some variations, distally ( Fig. 10 View FIGURE 10 T). Exopod of uropod extends 38– 44% its length beyond telson. Telson trapezoidal (apart from cleft), its maximum width near basis is 2.1–2.7 times that at apex ( Fig. 10 View FIGURE 10 U), its length is 1.9–2.5 times maximum width or 1.1–1.4 times length of sixth pleonite (measured along dorsal midline).
Statoliths ( Fig. 11 View FIGURE 11 A). The vaterite statoliths resemble ventrally flattened or truncated spheres, very similar to those of P. helleri ( Fig. 4 View FIGURE 4 in Ariani et al. 1993) and P. l a cu s t r i s ( Figs 4 View FIGURE 4 A, 6M in Wittmann et al. 1993); diameter 110–178 µm; statolith formula 2 + 3 + 1 + (5–9) + (11–16) = 22–31. Ambitus and tegmen with moruloid appearance, their surface resembling a wickerwork of lamellae. Such arrangements of lamellae often and so far exclusively found in vaterite statoliths. Ventrally the mantle does not cover the central part of the fundus, where a conical hole is typically visible (as in Fig. 11 View FIGURE 11 A).
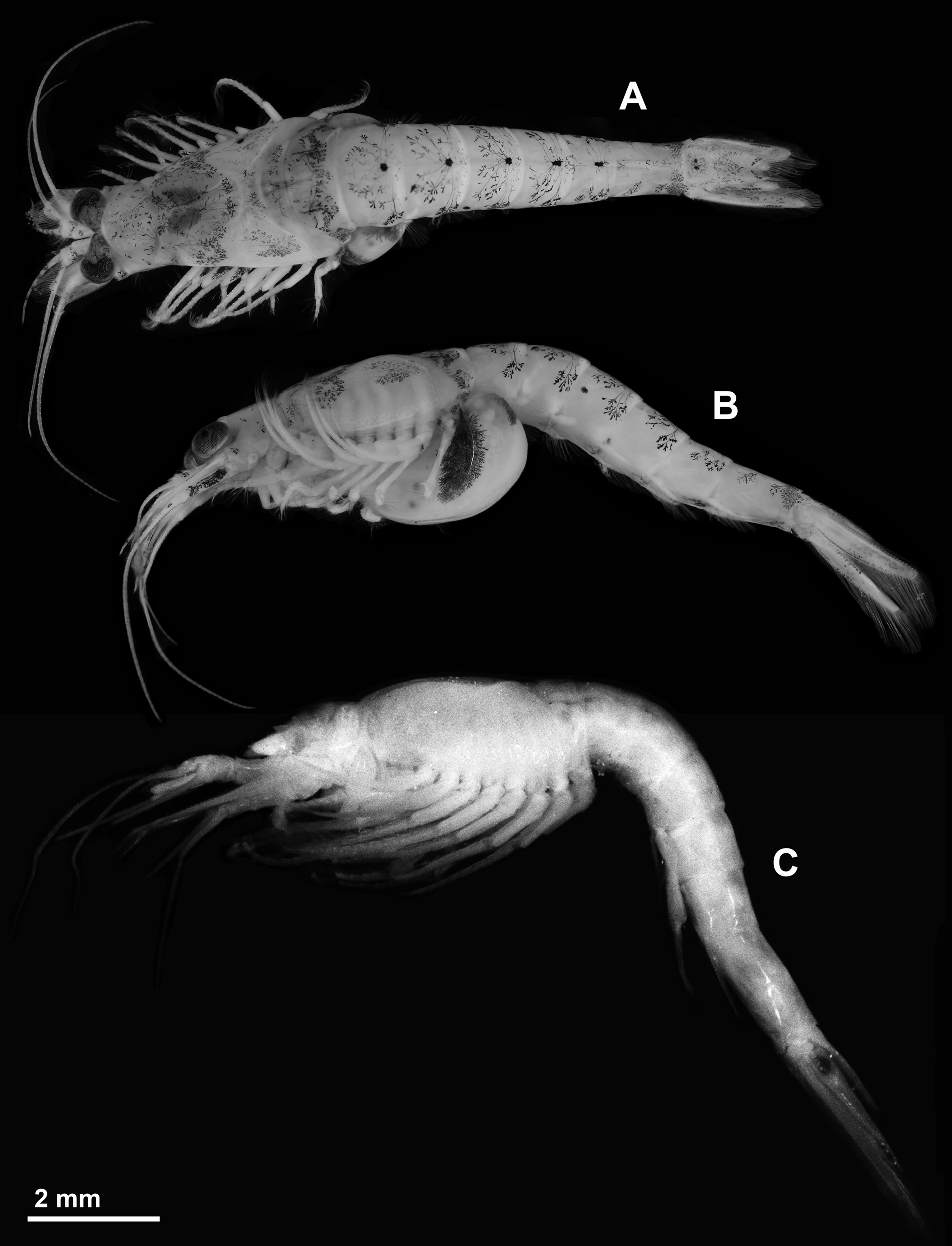
Colour. Not considering their less slender body and the larger eyes, freshly caught living specimens of the new species greatly resemble Diamysis mesohalobia mesohalobia Ariani & Wittmann, 2000 (colour photo in Ariani & Wittmann (2000): Fig. 2 View FIGURE 2 F) from Fiume Piccolo (see below). General appearance yellowish brown to dark brown depending on chromatophore expansion. Rich with chromatophores all over the body and appendages. The arborescent chromatophores give brown, yellowish, and to a lesser extent also rose markings. Mainly the brown components are preserved in the ethanol-fixed specimen shown in Fig. 7 View FIGURE 7 A, B.
Comparison. The new species is most similar to P. agigensis known from brackish and marine waters of the Black, Marmora, and Aegean Seas, with which it shares a short terminal lobe of the antennal scale, heavily armed inner paradactylar seta on thoracic endopod 3, and the few spines on endopod of uropod. It shares scale-like hairs on rounded lobes of anterior (2–5) male thoracic sternites with P. intermedia , P. ullskyi , P. kessleri sarsi , and P. bakuensis (for lobe morphology in Paramysis see Wittmann & Ariani (2011)). In contrast, there are no hairs on the rounded lobes of P. h e l l er i, P. festae , and P. nouveli , or on the less well-rounded lobes of P. k ro y er i. There are also smooth, but smaller, less distinct lobes in P. arenosa ( G. O. Sars, 1877) . The respective lobes are not rounded but subtriangular to sickle-shaped at least in large males of P. a gi ge n s i s, P. pontica , and P. bacescoi .
Additional differences of P. a dr i at i c a from its most similar (Ponto-)Mediterranean congenerics: from P. he l l e r i by shallower telson cleft (15–24% telson length in the latter), stouter antennal scale (2.8–3.9 times as long as wide in P. he l l e r i), strong, less numerous secondary spinules (about 20 thin distal setules in P. he l l e r i) on the paradactylar setae in endopods 3–8, reminiscent of those of P. agigensis , and by fewer spines on the endopod of uropods (7–13 in P. helleri ), also like in P. ag i g e n s i s. The new species differs from P. f e s t a e by smaller eyes, a longer antennal scale, and a greater number of laminae in the telson cleft. It differs from P. agigensis by a shallower telson cleft (15–19% telson length in P. agigensis ) not or barely reaching up to the penultimate lateral spines (almost reaching the antepenultimate spines in P. agigensis ), by slightly stouter antennal scale (2.8–3.0 times as long as wide in P. agigensis ), but slightly longer apical part of the antennal scale (16–18% scale length in P. a gi ge n s i s), and by the presence of 4–6 short, barbed setae on caudolateral blade of penis (1–3 long, smooth setae in P. agigensis ).
Pleopods of two adult males each from shallow brackish waters at sandy beaches were dissected for P. he l l e r i from the North Adriatic Sea ( Spiaggia die Ponente , Caorle, Italy, 45.5814N 012.8513E, S = 17, leg. KJW) and P. agigensis from the Sea of Marmora (Büyükada Islands, Turkey, 40.8594N 029.1126E, again S = 17, leg. KJW). The fourth pleopod of both species differed from that in P. adriatica by two-segmented (versus unsegmented) endopod and 7-segmented (versus 6-segmented) exopod, the latter showing a weaker, less strongly ridge-like differentiation at its upper surface near the end of its second segment (corresponding to the basal segment in P. adriatica ). GoogleMaps




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Mysinae |
|
Tribe |
Mysini |
|
Genus |
|
|
SubGenus |
Longidentia |
Paramysis (Longidentia) adriatica
| Wittmann, Karl J., Ariani, Antonio P. & Daneliya, Mikhail 2016 |
Paramysis helleri:
| Holmquist 1955 |
Mysis oculata relicta:
| Stammer 1932 |