
Paraleyrodes perseae (Quaintance)
|
publication ID |
https://doi.org/ 10.5281/zenodo.171844 |
|
DOI |
https://doi.org/10.5281/zenodo.6263613 |
|
persistent identifier |
https://treatment.plazi.org/id/255987D0-FFB2-7E1D-FEA7-FE35FB7BFD61 |
|
treatment provided by |
Plazi |
|
scientific name |
Paraleyrodes perseae (Quaintance) |
| status |
|
Paraleyrodes perseae (Quaintance) View in CoL
( Figs 5–10 View FIGURES 5 – 7 View FIGURES 8 – 16 )
Aleurodes perseae Quaintance, 1900: 32 –33. Lectotype, USA (Florida), here designated. Aleurodicus perseae (Quaintance) Cockerell, 1903: 663 .
Paraleyrodes perseae ( Quaintance) Quaintance, 1909: 170 View in CoL –173; Quaintance & Baker, 1913: 82 – 83.
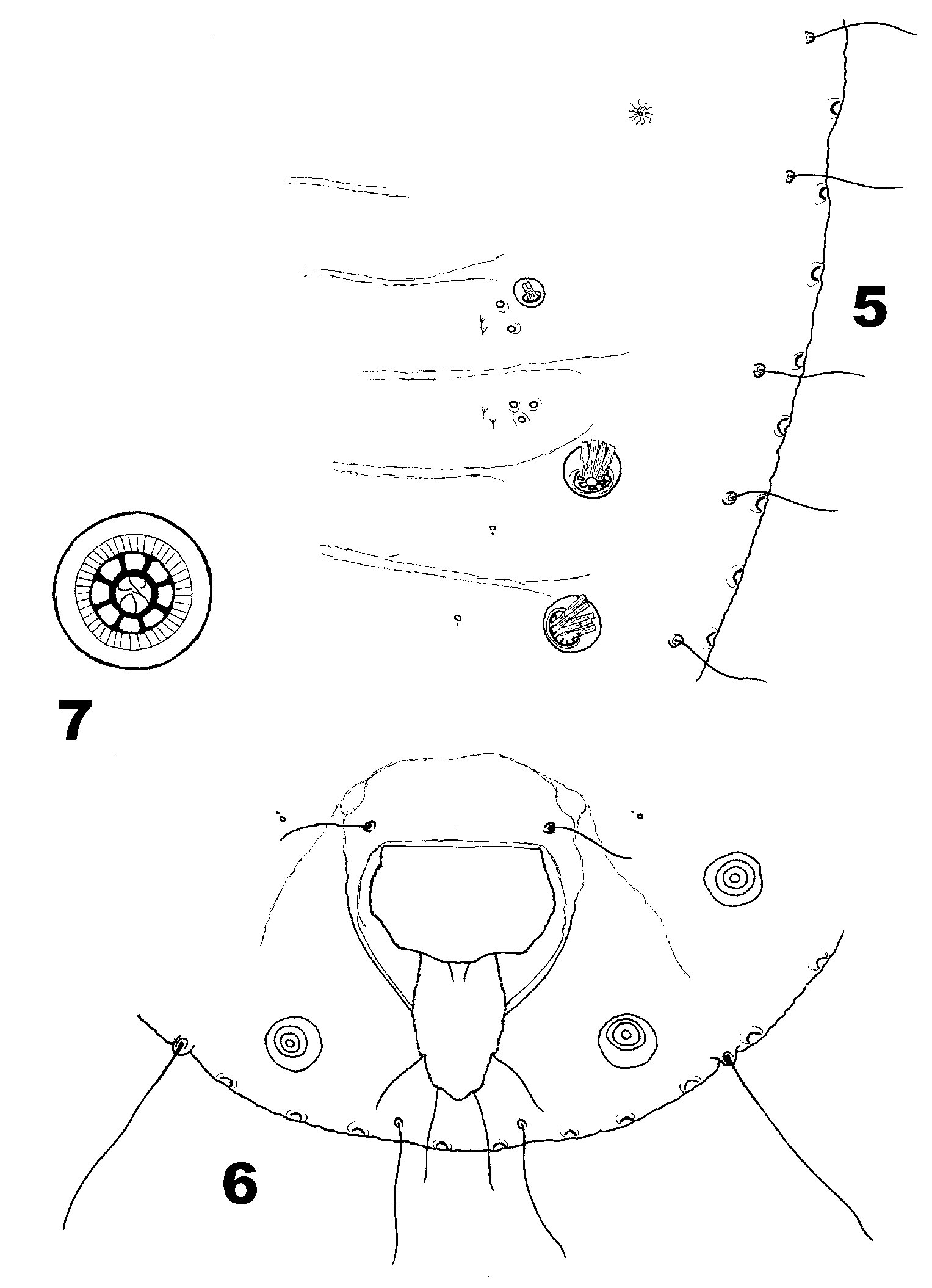
PUPARIUM (n=20). 0.62–0.90 mm long, outline oval, margin almost smooth. One pair of large compound pores ( Fig. 7 View FIGURES 5 – 7 ) present on cephalic segment and each of abdominal segments V–VIII, all similar in size with overall diameter 20–36 (usually 24–32) m; ring of spinneret cells 10–18 (usually 12–16) m in diameter, with 4–8 cells ( Fig. 7 View FIGURES 5 – 7 ), each with an elongate spline issuing from it ( Fig. 5 View FIGURES 5 – 7 ); between ring of spinneret cells and boundary of compound pore is a narrow annulus with much finer cells; inner lumen of compound pore (when detail can be seen) divided into irregular segments ( Fig. 7 View FIGURES 5 – 7 ). A single pair of small compound pores present, on abdominal segment III, 12–16 m in diameter, about the same size as the thoracic cicatrices ( Fig. 5 View FIGURES 5 – 7 ); the pair of small compound pores usually present on abdominal segment IV of Paraleyrodes View in CoL puparia is absent. Subdorsum sparsely provided with small pore/porette geminate pairs, usually a single pairing to either side of abdominal segments V–VII (but not VIII) ( Figs 5–6 View FIGURES 5 – 7 ) and most cephalothoracic segments; abdominal segments III and IV each with a subdorsal cluster of 2–5 slightly larger “bright” pores and (mesad) 1–2 tiny spines ( Fig. 5 View FIGURES 5 – 7 ). Outer submargin with a ring of widelyspaced small craterlike glands ( Figs 5–6 View FIGURES 5 – 7 ) between the ring of submarginal setal bases and puparial margin, each one similar in size to a setal base, about 2–4 per pair of setae. Vasiform orifice ( Fig. 6 View FIGURES 5 – 7 ) cordate, smooth; operculum as figured, with a pair of tiny setae near midpoint of posterior edge; lingula head as figured, slightly angular.
ADULT MALE (n=8). Body 0.90–1.00 mm long, genital segment 0.115–1.30 mm in maximum dimension, claspers 0.09–0.10 mm long, antennal flagellum 0.37–0.48 mm long. Aedeagal apex as shown in figures 8–10, with 4 ventrallydirected short “thorns” variably visible, depending on precise alignment on slide (lateral or oblique aspect is needed for diagnostic purposes). Vasiform orifice, operculum and lingula as shown, the lingula subtly lobulate ( Fig. 8 View FIGURES 8 – 16 ).
SPECIMEN DEPOSITORIES.
BMNH—The Natural History Museum, London SW7 5BD, U.K.
CDFA—California Department of Food & Agriculture, Sacramento, California 95832, U.S.A. USNM—U.S. Department of Agriculture, Beltsville, Maryland 20705, U.S. A [Sternorrhyncha collections of the United States National Museum of Natural History / Smithsonian Institution].
MATERIAL EXAMINED (arranged chronologically).
Lectotype puparium of Aleurodes perseae , USA—FLORIDA, Fort George, on Persea carolinensis , 22 iv.1880 (Dr R.S.Turner) ( USNM); paralectotypes, 1 puparium, 16 firstinstar larvae and egg remains, all on same slide as lectotype ( USNM); 1 parasitised puparium, 2 adult males, USA—FLORIDA, Orlando, on Citrus limon , 28 i. 1907 (Morrill, Quaintance collection #5204) ( USNM); 1 puparium, USA—FLORIDA, St. Augustine, on “bay” [preseumed to be Laurus nobilis ], 29 vii. 1910 (Back, Quaintance collection #20223) ( USNM); 4 puparia, USA—VIRGINIA, Vienna, no host data, 31 x. 1914 (Williams) ( BMNH); 2 puparia, USA—FLORIDA, Orlando, on Citrus aurantium , no collection date but registered BM 1915410 (Hargreaves) ( BMNH); 3 puparia, 3 thirdinstar larvae, USA—GEORGIA, Darien, on Citrus [satsuma], 24 xi. 1943 (Mayer #396) ( USNM); 5 puparia, 1 thirdinstar larva, USA—MISSOURI, St. Louis, on Persea pubsecens , 27 i. 1944 (Mayer) ( USNM); 2 puparia, USA—FLORIDA, Longwood, on Cinnamomum sp., 16 vi. 1960 (Youtsey, via Weems) ( USNM); 1 puparium, USA—FLORIDA, Lake Monroe, on Viburnum odoratissimum , 30 xii. 1963 (Youtsey) ( USNM); 4 puparia, 1 adult male, USA—FLORIDA, Tallahassee, on Anthurium stems and leaves, 15 iii. 1965 (Weems via Miller) ( USNM); 1 puparium, USA—FLORIDA, New Smyrna Beach, on Myrica cerifera , 09 iii. 1972 (Roberts) ( USNM); 4 puparia, USA—HAWAII, intercepted exPHILIPPINES, on Piper betle , 29 ii. 1980 (intercept #048458) ( USNM); 1 puparium, USA—ILLINOIS, intercepted exMEXICO, on Psidium guajava , 20 i. 1981 (intercept #3888) ( USNM); 3 puparia, VENEZUELA, Aragua, La Candelaria, on Persea americana , 1.iii.1982, (Arnal) ( USNM); 1 puparium, 3 thirdinstar larvae, USA—CALIFORNIA, intercepted exPHILIPPINES, on Psidium guajava , 10 vi. 1982 (intercept #31335) ( USNM); 7 puparia, 1 thirdinstar with developing puparium, 7 adult males, NICARAGUA, Rio San Juan / Rio Bartola confluence, near El Castillo, on Persea americana , 23 vi. 2004 (Martin #8080) ( BMNH). [Note—as discussed in the Introduction, no characters have been identified that allow adult females of Paraleyrodes to be determined to species: with the frequency of mixed populations occurring (see Conclusions, below), all females are regarded as only generically determined and are not listed here.]
The lectotype of Aleurodes perseae is one of two puparia on a slide bearing the type data (see above), is clearly indicated on the adjacent red USNM type label, and is the specimen with clearly developing adult eyes visible through the cuticle. The lectotype is the specimen in best condition, and the author considers that improvement through remounting, with proper maceration, would be possible in the future.
DISTRIBUTION. USA—Florida, Georgia, Missouri, Virginia; Neotropical Region—Mexico, Nicaragua, Venezuela; Austrooriental Region—[ Philippines]; Pacific Region—Hawaii.
HOST PLANT GENERA. Anthurium , Cinnamomum , Citrus , Laurus , Myrica , Persea , Piper, Plumeria , Psidium , Viburnum .
DESCRIPTIVE CONFLICT—PUPARIUM. In the original description of Aleurodes perseae Quaintance (1900) stated that “On the dorsum are 5 pairs of cupshaped compound pores, 4 pairs on caudal third of case [posterior part of abdomen], and the fifth pair on cephalic segment…..”. However, the puparium was illustrated with five abdominal pairs of compound pores, the anteriormost abdominal pair being significantly smaller than all the others. The original account also described puparial secretions that were much more extensive than is usually seen amongst Paraleyrodes species: in a later paper Quaintance (1909) quoted the opinion of Dr H.W. Morrill, that puparia with such secretions had proved to be parasitised, and that is confirmed by the present author’s field observations of Paraleyrodes puparia ( Fig. 4 View FIGURES 3 – 4 ). This then raised the question of whether the originallydescribed form might have been atypical, due to parasitism, with normal specimens (not seen by Quaintance at that time) perhaps having the typical complement of seven pairs of compound pore pairs in total.
Quaintance (1909) redescribed Aleurodes perseae , and formally designated it as the type species of his new genus Paraleyrodes . In his “revised description” the compound pore complement did indeed rise from five pairs (described) and six pairs (illustrated) [in 1900] to seven pairs (both described and illustrated), this being the typical situation throughout the genus as now understood. This revised description was repeated by Quaintance & Baker (1913), but with fresh illustrations provided. Whereas the 1909 drawing was a fairly accurate representation of a typical Paraleyrodes puparium, the 1913 puparial figure showed all six abdominal compound pores to be the same size as each other, a characteristic of Metaleurodicus species that is never seen in Paraleyrodes .
Although puparia with only five pairs of abdominal compound pores were generally (and correctly, see Conclusions) being identified as P. perseae for many years, Paulson & Kumashiro (1985) stated that puparia of Hawaiian P. perseae bore six abdominal pairs, thus continuing the confusion over this species’ identity. However examination of material on behalf of the author (Bernarr Kumashiro, personal communication) indicates that the Hawaiian record is correct but that Paulson & Kumashiro’s description was not.
DESCRIPTIVE CONFLICT—ADULT MALE. In the original description Quaintance (1900) quoted Theodore Pergande’s very basic observations on an adult, but the sex was not noted and Quaintance stated that the single specimen was lost before he could examine it himself. Pergande’s colour observations do suggest that the lost specimen might indeed have belonged to Paraleyrodes , but nothing more can be deduced.
In his 1909 redescription, Quaintance did describe adults and spoke of Morrill having “been able to obtain the adult in quantity” and having “kindly furnished [Quaintance] with abundant specimens of all stages…..”. However, he described adults generally as having four antennal segments, which is a character of female Paraleyrodes only, and the caption of his illustration of such a female antenna simply says “antenna of adult”, suggesting that he did not study any male antennae at all. Quaintance clearly did study at least one male specimen, and the aedeagal apex was illustrated as a twopronged structure (reproduced here, Fig. 16 View FIGURES 8 – 16 ), but the lack of comment on the highly unusual and characteristic male antennae is remarkable. In Quaintance & Baker’s (1913) new illustrations, that accompanied the repeated 1909 revised description of “ perseae ”, the bifurcate male aedeagus seems to have been drawn from the same specimen figured in 1909, but the drawing is not quite identical; also a clearlyfemale antenna is again figured and captioned as though it could have belonged to either sex.
A single male in BMNH ( USA, Florida, Orlando, no host data, 12.xi.1914, Williams, Fig. 14 View FIGURES 8 – 16 ) has a bifurcate aedeagus, but with the prong bases not contiguous as illustrated in the 1909 and 1913 papers. Two further male specimens, in CDFA, ( USA, Florida, Broward Co., Davie, Citrus sp., 08.xii.1985, Dowell, Fig. 15 View FIGURES 8 – 16 ) each have a bifurcate aedegus, but with the prongs almost parallel and directed posteriorly, rather than laterally as in the Williams specimen. Either of these males may belong to the species which Quaintance and Quaintance & Baker thought was P. perseae , but both remain undescribed and unassociated with puparia. The aedeagus depicted in the 1909 and 1913 papers also bears some resemblance to P. a n c o r a Martin (2004) ( Figs 12–13 View FIGURES 8 – 16 ), described from material on avocado in Belize and Nicaragua; however, P. a n c o r a also features a dorsoapical aedeagal process not present in the Quaintance / Quaintance & Baker drawings, and has not yet been recorded from anywhere in the USA.
CONCLUSIONS. After so many years of uncertainty concerning the identity of P. perseae , recent observations have enabled a resolution of the problem, through examination of puparia, associated adult males, and puparia containing fully developed preemergence males. The 1900 puparial illustration is correct in its depiction of the compound pores, the “missing” pair of small compound pores is not an artifact of parasitism, and this confirms that P. perseae is the only known species of Paraleyrodes with a pair of very small compound pores on abdominal segment III, but none at all on segment IV. For the future, the distinctive puparia will probably allow accurate identification, even in the absence of confirmatory males. In practice, puparia have hitherto tended to be correctly identified, and it is the males that have not.
The aedeagal apex of male specimens recently collected in Nicaragua ( Figs 8–9 View FIGURES 8 – 16 ), and reared from puparia that match the lectotype puparium of Aleurodes perseae , is quite different in structure from that which Quaintance (1909), and subsequently Quaintance & Baker (1913), mistook for P. perseae . This finding, combined with the illustration of a puparium with six pairs of abdominal compound pores ( Quaintance, 1909), indicates that the redescription of “ P. perseae ” involved a different species entirely. Unfortunately, the detail of the 1909 description is still insufficient to lead to any conclusion about the species concerned.
The aedeagus of true P. perseae is illustrated here (probably for the first time) in figures 8–10. This should allow for confident identification of this species when puparia are also present. However, the male aedeagus of P. perseae is very similar to that of P. citricolus Costa Lima ( Fig. 11 View FIGURES 8 – 16 ), which typically exhibits six, somewhat digitiform, ventral processes but which otherwise shares the same lateral profile. That P. perseae and P. citricolus are particularly closely related is also indicated by the structure of the inner lumen of the large compound pores ( Fig. 7 View FIGURES 5 – 7 ), even though the large compound pores of P. citricolus are usually over twice the diameter of those of P. perseae .
A final observation is that this is the second example of a species of Paraleyrodes being incorrectly identified because described males were not associated with puparia. For many years, P. pseudonaranjae Martin (2001) was misidentified as P. naranjae Dozier (1927) , based on its puparia. However, the males in invasive populations did not match the two male syntypes from Dozier’s original sample of P. naranjae , and the invasive populations proved to be a different and undescribed species. The author’s own field experience indicates that, in their native habitat, puparia of Paraleyrodes species are frequently found with adults of different species feeding on the same individual plant, making the association of adults with puparia difficult. Conversely, invasive populations tend to be taxonomically useful, for they usually comprise only single species.


| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paraleyrodes perseae (Quaintance)
| Martin, Jon H. 2006 |
Paraleyrodes perseae ( Quaintance) Quaintance, 1909 : 170
| Quaintance 1913: 82 |
| Quaintance 1909: 170 |
Aleurodes perseae
| Cockerell 1903: 663 |
| Quaintance 1900: 32 |