
Nudiantennarius subteres (Smith and Radcliffe, in Radcliffe)
|
publication ID |
https://doi.org/ 10.1643/CI-17-651 |
|
DOI |
https://doi.org/10.5281/zenodo.6040701 |
|
persistent identifier |
https://treatment.plazi.org/id/910E87DB-FFEC-706B-FFE6-1FCBFBB52A4D |
|
treatment provided by |
Plazi |
|
scientific name |
Nudiantennarius subteres (Smith and Radcliffe, in Radcliffe) |
| status |
|
Nudiantennarius subteres (Smith and Radcliffe, in Radcliffe) View in CoL
Lembeh Frogfish
Figures 1–5 View Fig. 1 View Fig. 2 View Fig. 3 View Fig. 4 View Fig. 5
Antennarius subteres: Smith and Radcliffe, in Radcliffe, 1912:205 View in CoL , pl. 17, fig. 1 (original description, single specimen, holotype USNM 70268, Lingayen Gulf, west coast of Luzon, Philippines).
Nudiantennarius subteres: Schultz, 1957:66 View in CoL , pl. 1, fig. D (new combination; after Smith and Radcliffe, in Radcliffe, 1912). Pietsch, 1984:36 (genera of frogfishes). Pietsch and Grobecker, 1987:184, figs. 16C, 75–77, 129 (description, distribution, relationships; new records from Luzon and Ambon). Lindberg et al., 1997:213 (Sea of Japan). Pietsch, 1999:2015 (western central Pacific, in key). Pietsch, 2000:597 (South China Sea). Allen and Adrim, 2003:25 (Moluccas; Flores locality based on misidentification: NMV A.9676, 11 mm SL, is Antennarius hispidus View in CoL ). Kuiter and Debelius, 2007:117, color fig. (misidentification, after Allen and Adrim, 2003). Allen and Erdmann, 2012:155 (Indonesia). Arnold and Pietsch, 2012:128, fig. 1E (molecular phylogeny).
Antennarius View in CoL sp.: Allen et al., 2003:363, color fig. (‘‘Ocellated Frogfish,’’ ‘‘unidentified, possibly undescribed species known only from Lembeh Strait, Sulawesi’’).
Material examined.— Seven specimens, 17–64 mm: holotype of Antennarius subteres , USNM 70268 , 42 mm, Albatross station 5442, Lingayen Gulf, west coast of Luzon , Philippines, 16°30"36" "N, 120°11"06" "E, beam trawl, 82 m, on a bottom of coral sand, 10 May 1909. Additional material: BMNH 1866.8.14.108, 64 mm, cleared and stained (now badly degraded), locality and depth unknown, Damon; CAS 32765, 32 mm, Bataan, 11.3 km west of Talaga, Luzon Island, Philippines, 13.7364°N, 120.8314°E, between 64 and 90 m, J. E. Norton, 1 June 1966; CBG (Center for Biodiversity Genomics, University of Guelph), Barcode of Life voucher 13028, misidentified as Antennarius nummifer (located by blasting the COI gene sequenced from the UW specimens), 42 mm, near Manila, Luzon, Philippines, 14.246°N, 120.479°E, additional data unknown; UW 117643, 30.5 mm, male (?), aquarium specimen, locality unknown; UW 119524, 38 mm, female with scrolled ovaries each containing hundreds of tiny eggs, Secret Bay, Gilimanuk, Bali, via Golden Ina Imports, additional data unknown; ZMUC P922045, 17 mm, Ambon Island, Moluccas, 128 m, Mortensen, 25 February 1922.
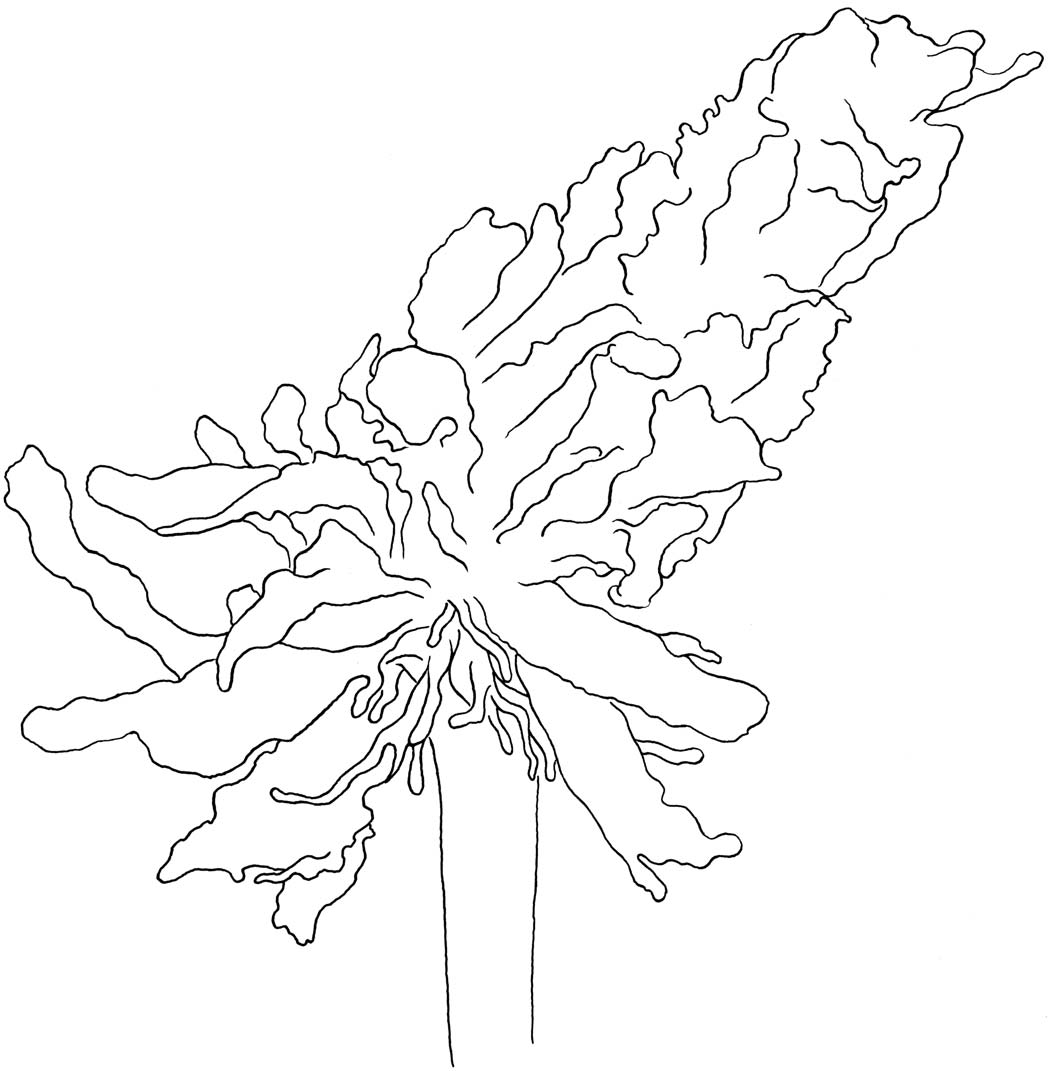
Diagnosis.— Nudiantennarius is unique among antennariids in having the following combination of character states: dermal spinules reduced, skin only partially covered with bifurcate dermal spinules, body often appearing naked, length of spines of each spinule not more than twice the distance between tips of spines ( Pietsch and Grobecker, 1987:fig. 16C); esca distinct ( Fig. 2 View Fig. 2 ); illicium naked, without dermal spinules, about half length of second dorsal-fin spine; second dorsal-fin spine unusually long, narrow, without posterior membrane; pectoral-fin lobe narrow, somewhat detached from side of body; caudal peduncle present, the membranous posteriormost margin of soft-dorsal and anal fins attached to body distinctly anterior to base of outermost rays of caudal fin; all rays of caudal fin usually bifurcate (outermost caudalfin rays simple, 7 innermost bifurcate in UW 117643 and CBG 13028); endopterygoid present; pharyngobranchial I present; epural present; pseudobranch absent; swimbladder present; dorsal-fin rays 12; anal-fin rays 7; pectoral-fin rays 9; pelvic-fin rays 5, all simple (posteriormost ray not bifurcated as erroneously stated by Arnold and Pietsch, 2012); membranes between rays of paired fins deeply incised ( Pietsch and Grobecker, 1987:fig. 77); one or more large basidorsal ocelli usually present.
Description.— Dermal spinules greatly reduced, evident on second and third dorsal-fin spines, anteriormost dorsal-fin rays, on snout and dorsal portion of head, often on chin, coverage sometimes extending to pectoral lobe, but elsewhere few and scattered, usually difficult to detect without microscopic aid; esca a rounded clump of folded tissue (in smallest specimen examined) or an oval-shaped tuft of short, more or less flattened appendages, sometimes with a few short filaments (in larger specimens; Fig. 2 View Fig. 2 ), usually directed anteriorly and often forming an acute angle with the illicium; length of esca 3.9–7.2% SL; illicium, when laid back onto head, fitting into a tiny, narrow groove situated alongside of second dorsal-fin spine, tip of illicium (esca) coming to lie within a shallow depression between second and third dorsal-fin spines, esca probably capable of being covered and protected by second dorsal-fin spine when spine is fully depressed; illicium about half length of second dorsalfin spine, 5.9–11.3% SL; anterior end of pterygiophore of illicium terminating distinctly posterior to symphysis of upper jaw; illicium and second dorsal-fin spine relatively closely spaced on pterygiophore, distance between bases of juveniles and that the adults, which are rarely seen, exist at much greater depths (Vincent Chalias, pers. comm., 11 April 2017). This notion is contradicted, however, by observations of small females through multiple reproductive cycles; in several cases, a heavily gravid female, no more than about 15 mm, has been observed in the company of two slightly smaller individuals, apparently males, foretelling a reproductive event (Daniel Geary, pers. comm., 4 May 2017).
spines less than 5% SL; second dorsal-fin spine long, narrow, straight to slightly curved posteriorly, not connected to head by membrane, sometimes with numerous slender, cutaneous filaments; length of second dorsal-fin spine 17.4–28.1% SL; third dorsal-fin spine curved posteriorly, connected to head by thick membrane, length 23.0–31.3% SL; eye diameter 5.5– 11.3% SL; only distal tip (about 20–25% of length) of maxilla tucked beneath folds of skin; scattered cutaneous appendages often present on head (especially on chin), body, and second and third dorsal-fin spines; epibranchial I toothless; ceratobranchial I toothless; vertebrae 19, caudal centra 14; dorsalfin rays 12, posteriormost 2–5 bifurcate; anal-fin rays 7, all bifurcate; pectoral-fin rays 9, all simple; all rays of pelvic fin simple; distal third to half of pectoral- and pelvic-fin rays free, not connected by membrane ( Fig. 1 View Fig. 1 ).
Coloration.— In life, highly variable: typically, overall dark purplish brown, chocolate brown, to black, with a large brown to black basidorsal ocellus, usually surrounded by a light brown, yellow, orange, or red ring; similar but much smaller ocelli occasionally present on membrane behind third dorsal-fin spine, anterior half of soft dorsal fin, and upper margin of caudal peduncle; tiny scattered white spots often present on head, body, and fins, especially on caudal fin; illicium, esca, and all or part of second dorsal-fin spine light brown to white; free distal tips of paired fins light brown to almost white. Variations on this general theme are seemingly endless: background coloration may range from pink, red, orange, yellow, green, or gray to off-white; head, body, and fins often monochromatic, but sometimes mottled or covered with small dark, close-set spots; ring of ocellus sometimes surrounded by a series of dark close-set spots; ocellus sometimes absent; illicium without banding; small dark bars of pigment often radiating from eye ( Figs. 3 View Fig. 3 , 4 View Fig. 4 ).
Size.— Nudiantennarius subteres is apparently a relatively small frogfish. Six of the seven known preserved specimens range from 30.5–64 mm, but by far most estimates of uncollected individuals are considerably smaller, no more than about 22 mm. Accordingly, some observers have hypothesized that the numerous sightings of living specimens are based on Distribution.— Originally described from a single specimen collected in Lingayen Gulf, on the west coast of Luzon Island, Philippines, the additional preserved specimens are from Manila and off Bataan, west of Talaga, Luzon Island; Secret Bay, Gilimanuk, on the west end of Bali; and Ambon, Moluccas Islands. Photographs, however, expand the distribution to include Lembeh Strait and Bangka Island, North Sulawesi; Pantar and Alor islands in the Alor Archipelago; and Dauin and Dumaguete, Negros Island, Philippines. Early collection data indicate a deep-water existence, at depths between 64 and 90 m (CAS 32765), 82 m (the holotype, USNM 70268), and 128 m (ZMUC P922045)—the reason why Pietsch and Grobecker (1987) called it the ‘‘Deepwater Frogfish’’—but numerous recent observations by divers indicate a much shallower existence, from 3–30 m, although some hypothesize that most observations are based on juveniles, while adults occur at greater depths (Vincent Chalias, pers. comm., 11 April 2017).
Habitat.— Lembeh Strait is a narrow body of water, approximately 1.5 km wide and several kilometers long, between Lembeh Island and the northeastern corner of Sulawesi, Indonesia. Most dive sites are located along the Sulawesi side and close to shore; depths vary, but most range from 3–9 m, with no sites deeper than about 18 m. There are large amounts of tree litter that have fallen into the water in the shallows, and a lot of trash, rubbish, and human refuse of all kinds, at many locations, especially those near native villages and towns. Bottoms on which the Lembeh Frogfish have been observed are typically brown or black sand, silt, or mud, with some soft corals, gorgonians, and sponges, but very little hard coral (David J. Hall, pers. comm., 7 April 2017). It has also been observed on pier pilings and occasionally among small patches of filamentous and leafy algae (Scott W. Michael, pers. comm., 22 March 2017).
The Pantar dive site, located on the southeast margin of the island in the Alor Archipelago, Indonesia, where David Hall photographed the Lembeh Frogfish in 2006 (pers. comm., 7 and 9 April 2017), is a much smaller area than Lembeh Strait—a small bay adjacent to a small native village. Most of the diving was no deeper than 6 m and the bottom was largely made up of very coarse sand or fine coral rubble, with some hard corals and gorgonians, but, unlike Lembeh, no rubbish strewn over the bottom. There were numerous small cephalopods, many shrimps, crabs, other crustaceans, and lots of fishes, especially juveniles: small shark species, burrowing snake eels, two different species of Rhinopias (R. eschmeyeri and R. frondosa ), and several other species of scorpionfishes and waspfishes. Only two species of frogfishes were observed, Antennarius striatus and a single individual of the Lembeh Frogfish.
Most of the dive sites in Ambon Bay where the Lembeh Frogfish has been observed offer typical ‘‘muck diving ’ habitat—coarse sand- and rubble-covered slopes, with random solitary and encrusting sponges, hydroids, mixed with lots of human refuse, the latter more or less covered with organic growth, which the animals use for shelter (Linda Ianniello, pers. comm., 5 June 2017). At Bali, the species occurs most often on black sandy slopes on the northeast margin of the island, and most commonly seen during night dives (Vincent Chalias, pers. comm., 11 April 2017). At Dauin on Negros Island, Philippines, the habitat is similar: darkcolored individuals are most commonly found on coarse sand or gravel, often within patches of green algae, in 4–20 meters; lighter, more colorful individuals are usually found associated with small, similarly colored sponges, at somewhat greater depths, 12–30 m (Daniel Geary, pers. comm., 4 May 2017).
Relationships.— A combination of morphological and meristic characters, including the presence of a endopterygoid and an epural, 19 vertebrae, and double scroll-shaped ovaries ( Pietsch and Grobecker, 1987; Arnold and Pietsch, 2012), indicates that N. subteres is a member of the frogfish subfamily Antennariinae , as diagnosed by Arnold et al. (2014). Within the Antennariinae , both morphological and molecular analyses indicate a close relationship with the Sargassumfish, Histrio histrio ( Fig. 5 View Fig. 5 ). Shared morphological features include a short illicium, no more than half the length of the second dorsal-fin spine; the second dorsal-fin spine unusually long and narrow, without a posterior membrane; the illicium and second dorsal-fin spine closely spaced, the former appearing to emerge from the base of the latter; all rays of the pelvic fin simple; and greatly reduced dermal spinules. In addition, the pectoral-fin lobe of both species is detached from the side of the body, partially so in N. subteres but free for most of its length in H. histrio ( Pietsch and Grobecker, 1987) . Finally, a Bayesian analysis of COI sequences recovered N. subteres as sister to H. histrio , with a posterior probability of 1.0 ( Fig. 5 View Fig. 5 ). Genetic distances between N. subteres and H. histrio differed by 4.8–5.5%, and 1.1% between the two specimens of N. subteres .
Reproduction.— The presence of double scroll-shaped ovaries in Nudiantennarius , hypothesized by Arnold and Pietsch (2012) and now verified in UW 119524 (apparently the only female among the known preserved specimens of this species), corroborates placement of the genus within the Antennariinae and suggests that it undergoes a distinct pelagic larval stage before metamorphosis to the adult form, in contrast to members of the Histiophryninae , which undergo direct development and employ various modes of parental care ( Arnold et al., 2014).
DISCUSSION
Since its original description more than a century ago, Nudiantennarius subteres has remained somewhat of an enigma. Although Pietsch and Grobecker (1987) were able to assign three additional specimens, it has never been well understood. The absence of field observations and its rarity in collections has generally been attributed to a deep-water distribution, all known specimens at the time having been taken below 60 m, well beyond standard SCUBA depth. The ‘‘Lembeh Frogfish,’’ on the other hand, has also posed an enigma. Its distinctive morphology has been recognized among members of the dive community for almost two decades and among professional ichthyologists at least since the early 2000s (see Allen et al., 2003). It is therefore a bit of a mystery that no formal description and scientific name has been applied to it. The many images attributable to this species (posted on the internet and sent to us by many underwater photographers; see Acknowledgments) have been variously identified as Antennarius biocellatus , Antennatus nummifer, Antennatus dorehensis, and Antennatus rosaceus, but clearly it cannot be assigned to any of these species—the sum of its characters points to Nudiantennarius subteres . The combination of reduced dermal spinules, unique escal morphology, short illicium, and long second dorsal-fin spine, unattached to the head by a membrane, is enough to separate the species from all other frogfishes, except Histrio histrio (see above)—the latter remains unique in having exceptionally large pelvic fins and a pseudopelagic lifestyle in floating Sargassum .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nudiantennarius subteres (Smith and Radcliffe, in Radcliffe)
| Theodore W. Pietsch & Rachel J. Arnold 2017 |
Nudiantennarius subteres
| : Schultz 1957: 66 |
Antennarius subteres
| : Smith and Radcliffe, in Radcliffe 1912: 205 |