
Neocyclotus dysoni, AMBIGUUM
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00058.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B6B923-EE31-FFEB-8FEC-62343F0C6C86 |
|
treatment provided by |
Carolina |
|
scientific name |
Neocyclotus dysoni |
| status |
|
NEOCYCLOTUS DYSONI AMBIGUUM View in CoL (MARTENS 1890)
Material examined
Yucatan, Mexico ( USNM 890933 About USNM ) .
External anatomy and mantle cavity
Foot broad with narrow opening of anterior pedal gland. Operculum with outer calcareous layer and chitinous basal plate. Pallial roof well vascularized, ctenidium absent. Hypobranchial gland forming thick pad, narrowing anteriorly. Hypobranchial gland composed of subepithelial gland cells penetrated by ciliated ducts discharging to ciliated excretory groove. No differentiation of hypobranchial gland into two distinct regions.
Reproductive system
Gonad dorsally overlying digestive gland, extending to posterior end of midgut. Gonopericardial connection absent. Oviduct extending anteriorly past proximal tip of glandular pallial oviduct, looping back upon itself, broadening into lobulate seminal receptacle; receptacle lying along left, ventral wall of glandular oviduct bearing orientated sperm. Oviduct extending anteriorly from receptacle, sending off dorsal branch to large, saculate bursa copulatrix lying above receptacle, containing unorientated sperm. Oviduct continuing forward, gradually increasing in diameter and becoming more glandular, closely appressed to ventral side of glandular oviduct. Oviduct, opening to capsule gland short distance back from genital aperture.
Pallial oviduct partially embedded in viscera behind base of mantle cavity, extending to mantle edge. Albumen gland forming proximal end of glandular oviduct, alongside seminal receptacle and bursa, with gradual transition anteriorly to cells comprising capsule gland. Lumen of pallial oviduct divided longitudinally forming two chambers, more or less dorsal and ventral, communicating posteriorly via short ventro-lateral slit extending from bursa to near base of pallial oviduct. Dorsal chamber surrounding ventral chamber dorsally and on left. Oviduct opening to ventral chamber. Ventral chamber ending blindly anteriorly, becoming highly complexly folded posteriorly. Dorsal chamber bearing genital aperture at anterior end, with simpler internal structure, becoming less glandular posteriorly.
Seminal vesicle derived from vas deferens absent. Closed prostate lying along base of mantle cavity. Seminal groove leading from distal end of prostate to penis lying on side of head in centre of nape. Seminal groove secondarily closed with superficial groove overlying the duct.
Alimentary system
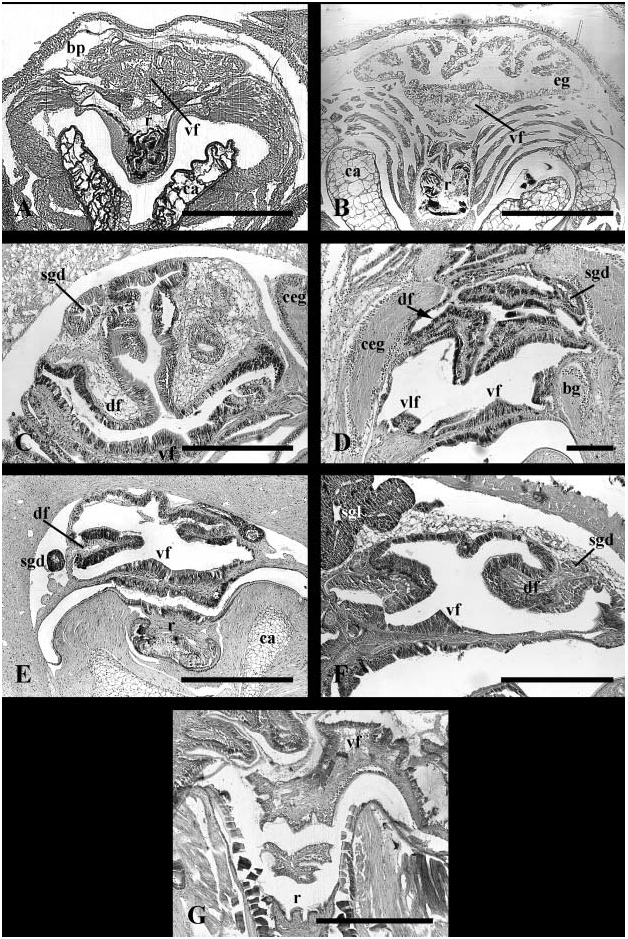
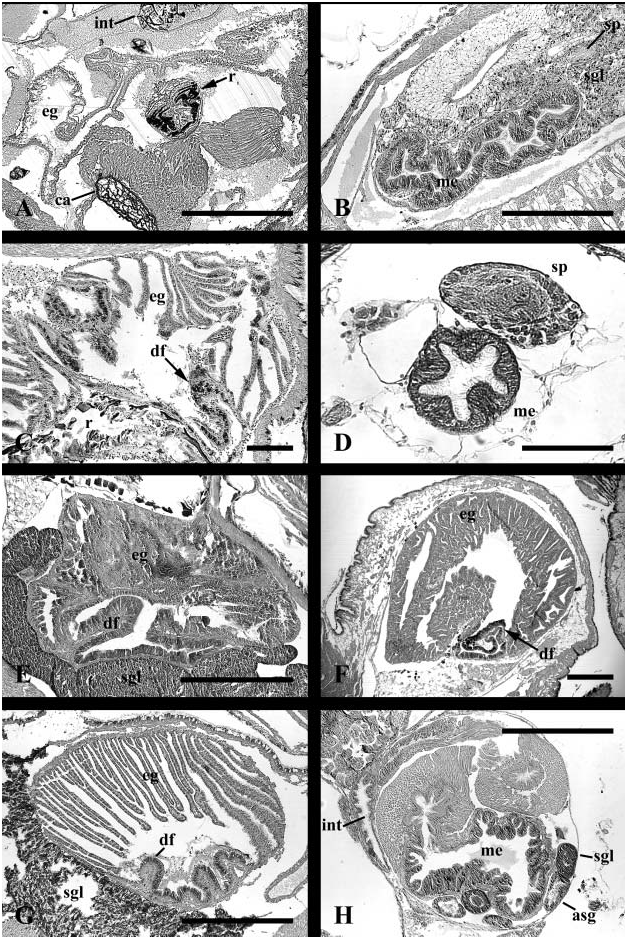
Foregut. Transverse mouth opening at tip of short snout to large buccal mass ( Fig. 6A View Figure 6 ). Radula taenioglossate. Buccal cavity walls bearing pair of prominent jaws with saw-like teeth, lying well forward of dorsal folds. Jaw composed of outer homogeneous layer and inner rod layer ( Fig. 5A, j View Figure 5 ). Subradular organ present ( Figs 4B View Figure 4 , 5A View Figure 5 , sro). Organ long and well developed with shape of flattened cone projecting between jaws into mouth opening. Shallow horizontal clefts marking sides of organ. Short ventral fold present, beginning in buccal cavity ( Fig. 8C View Figure 8 , vf). Ventral trough replacing ventral fold, flanked by two ventro-lateral folds ( Fig. 9C View Figure 9 , vlf) running along inner edges of ducts of buccal pouches (bp). Pouches short, broad ( Fig. 6A View Figure 6 , bp); pouch ducts lined with basophilic epithelium. Salivary gland ducts (sgd) opening dorso-laterally, under cerebral commissure. Salivary glands massive, overlying portions of mid- and posterior oesophagus to back of cephalic haemocoel. Mid-oesophageal epithelium uniformly folded after separation of pouches, with broad, flattened and longitudinally folded lumen; septate oesophageal gland absent ( Fig. 11B View Figure 11 , me).
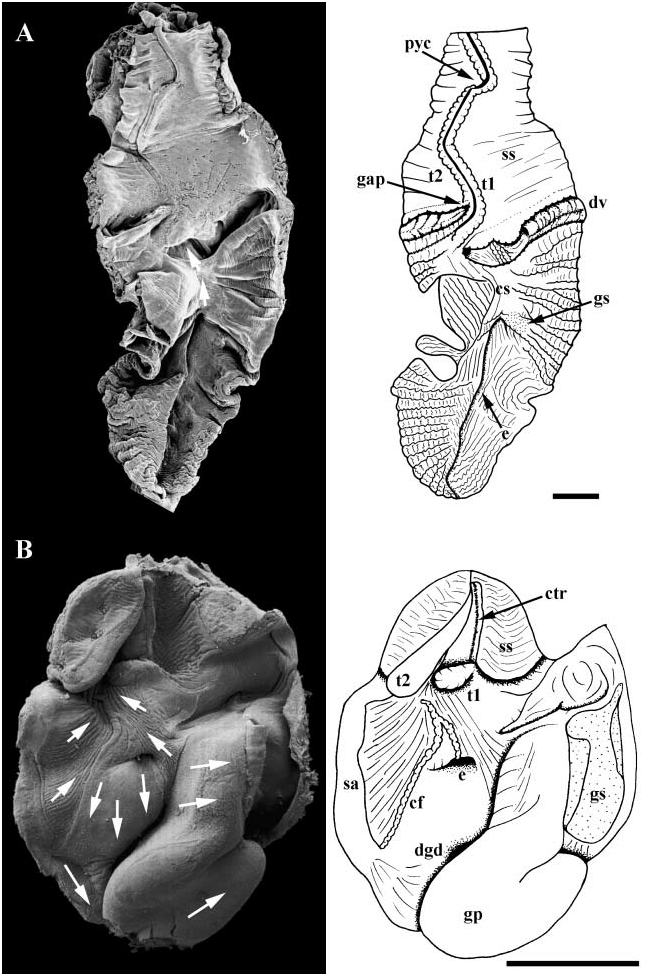
Midgut. Posterior oesophagus opening to left limb of U-shaped gastric chamber ( Fig. 13A View Figure 13 ). Distal portion of midgut almost completely lined by cuticle, leaving only a ventral ciliated strip (cs). Ciliary currents flowing anteriorly within ciliated zone. Cuticle elaborated into small gastric shield (gs) ventrally on right, adjacent to oesophageal opening (e). Large groove horizontally subdividing gastric lumen posterior to style sac. Deep, rounded posterior portion of groove forming vestibule receiving numerous digestive gland ducts (dv). Anterior tip ending in blind pouch (gap) next to intestinal groove; no secretion observed emanating from pouch. Shallow intestinal groove bounded by two low typhlosoles (t1, t2). Intestinal groove turning briefly to right at distal end of style sac. Transverse segment of intestinal groove bounding mouth of single pyloric caecum (pyc); caecum extending short distance posteriorly under style sac. Style sac epithelium smooth and unfolded, bearing well differentiated cilia. Raised ciliary tract on major typhlosole and crystalline style absent.
Hindgut. Intestine exiting style sac, extending short distance posteriorly alongside style sac, completing three full loops embedded in digestive gland.
Reno-pericardial system
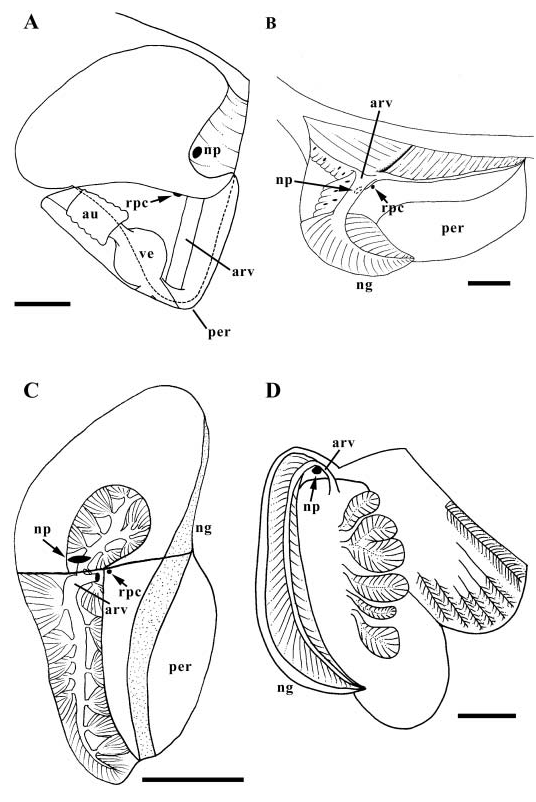
Kidney and pericardium lying within pallial roof ( Fig. 22A View Figure 22 ). Kidney large, overhanging pericardium; thick mass of excretory tubules almost completely filling lumen, roughly divided into several lobes. Nephridial gland absent. Nephropore (np) opening posteriorly to narrow vertical extension of mantle cavity behind kidney. Afferent renal vessel extending short distance back from cephalopedal sinus, turning dorsally, running along back wall of pericardium (per) to enter kidney from below, near nephropore. Afferent renal vessel entering kidney lobe overhanging pericardium. Reno-pericardial canal (rpc) opening dorsally near afferent renal vessel. Pericardial lumen narrow, extending short distance posterior to kidney.
Nervous system and sensory structures
Nervous system hypoathroid, right zygoneurous, left dialyneurous ( Fig. 24A View Figure 24 ). Circum-oesophageal nerve ring surrounding anterior buccal mass at level of ocular peduncles ( Fig. 6A View Figure 6 , nr). Labial commissure absent. Single, thick tentacular nerve innervating tentacle (tn). Pallial nerve arising from left pleural ganglion bearing small ganglion with several small branches, lying at base of pallial roof (osg). Pedal ganglia ( Fig. 24A View Figure 24 , pdg) connected by one commissure with two cross connections between pedal cords (pc). Statocysts (sc) with numerous statoconia on postero-dorsal surface of pedal ganglia. Buccal ganglia ( Fig. 6A View Figure 6 , bg) lying behind circum-oesophageal nerve ring at base of buccal cavity, just above buccal pouch ducts. Long connective leading to supra-oesophageal ganglion (sp) lying near back of cephalic haemocoel. Paired visceral ganglia (vg) lying under kidney, to right of pericardium, within cephalopedal sinus. Visceral commissure double. Osphradium absent. Eyes on tips of short ocular peduncles.
Remarks
Thompson (1969) described the reproductive anatomy of this species. While the gross morphology of that description is essentially identical to the one provided here, the interpretation of homologies is not. Thompson described the oviduct as persisting anteriorly along the distal end of glandular oviduct, with a short recurvent segment forming an albumen gland. However, as described above, the simple loop of the oviduct forms a seminal receptacle containing orientated sperm. Thompson also described a large, well-developed seminal receptacle closely appressed to the glandular oviduct, communicating with the oviduct distal to albumen gland. A bursa copulatrix was cited as lacking. However, the structure identified as the receptacle, is, in fact, a large bursa copulatrix with unorientated sperm. The glandular pallial oviduct was described as comprising only a capsule gland. However, the glandular pallial oviduct comprises both an albumen gland and a capsule gland. While not externally separable, these glands are distinguishable histologically. The description of male reproductive anatomy presented above is congruent with that of Thompson.
Discussion
There are few comprehensive anatomical studies of cyclophorids. Much of available comparative data concerns their reproductive anatomy, with only scattered accounts of nervous, alimentary and reno-pericardial anatomy. However, it seems clear that all cyclophorids share a unique combination of modifications to the mantle cavity. The pallial roof forms a highly vascularized respiratory surface ( Kasinathan, 1975). The ctenidium and osphradium have been lost, and the kidney, while lacking the ureter found in some terrestrial forms, has been displaced anteriorly and occupies a position within the pallial roof ( Tielecke, 1940; Andrews & Little, 1972; Kasinathan, 1975). The hypobranchial gland is modified into a thick pad penetrated by ducts and appears to have become an accessory excretory organ ( Andrews & Little, 1972). The size of the gland varies greatly among cyclophorid species, and may be differentiated into two distinct regions ( Thompson, 1969; Andrews & Little, 1972). However, the hypobranchial gland of Murdochia pallidum has been described as similar histologically to other marine ‘prosobranchs’ ( Morton, 1952a).
Descriptions of the reproductive system account for the majority of comparative studies among cyclophorids (e.g. Weber, 1924; Tielecke, 1940; Morton, 1952a; Berry, 1964; Thompson, 1969; Kasinathan, 1975; Jonges, 1980). The female is typically characterized by a pallial gonoduct divided into albumen and capsule glands, with a bursa and receptacle opening to the inner end of the albumen gland. In some species, the receptacle is not a separate pouch, but forms an enlarged section of the renal oviduct ( Morton, 1952a; present study). The pallial oviduct may open broadly to the mantle cavity ( Thompson, 1969; Kasinathan, 1975) or form a closed tube with a distal genital aperture ( Tielecke, 1940; Morton, 1952a; Berry, 1964; Thompson, 1969; Kasinathan, 1975). Male cyclophorids typically possess a prostate at the base of the mantle that may be open or closed ( Tielecke, 1940; Berry, 1964; Thompson, 1969; Kasinathan, 1975). A seminal groove, similarly open or closed, leads from the prostate to the penis lying behind the right cephalic tentacle or in the middle of the nape ( Tielecke, 1940; Morton, 1952a; Morrison, 1955; Berry, 1964; Thompson, 1969; Kasinathan, 1975). In some species, as in Neocyclotus dysoni ambiguum , the seminal groove may be secondarily coalesced by a raphe with a superficial groove overlying the duct ( Thompson, 1969).
A notable exception to this rather consistent picture is the reproductive anatomy of neotropical cyclophorids in the Aperostominae and Neocyclotinae ( Thompson, 1969). Thompson reported that all neotropical species studied lack a bursa copulatrix and possess an albumen gland formed by a simple loop in the oviduct, justifying unification of the two subfamilies in a new family, the Neocyclotidae . One of the species studied by Thompson included Neocyclotus dysoni ambiguum . The reanalysis of reproductive homologies for this species provided here are congruent with previous descriptions of cyclophorid reproductive anatomy and do not support recognition of a separate family for the neotropical genera.
There are virtually no published accounts of alimentary anatomy in cyclophorid species, and those that are available are contradictory. For example, buccal pouches may be present ( Amaudrut, 1898; Kasinathan, 1975) or absent ( Morton, 1952a). Amaudrut (1898) clearly figured a prominent subradular organ projecting between the jaws of Cyclophorus vulvulus . Morton (1952a) neither confirmed nor denied the presence of the subradular organ, but a schematic diagram of the foregut does not show a subradular organ. Similarly, Kasinathan (1975) did not report a subradular organ. The mid-oesophagus is narrow ( Kasinathan, 1975) or forms a voluminous crop ( Amaudrut, 1898).
Morton (1952a) described several features of the midgut of Murdochia pallidum congruent with the present description including a cuticularized epithelium with a ventral ciliated channel and the presence of a protostyle. However, Morton described no outpocketings of the gastric chamber nor style sac, the digestive gland is stated to open via a single duct near the style sac, and the intestine is reported as simple and S-shaped. Kasinathan (1975) described the gastric chamber as cuticularized only anterior to the oesophageal aperture; the presence of pyloric caecae or gastric pouches is neither confirmed nor denied, but a single digestive gland duct is reported in a position similar to that of the gastric pouch described here for Neocyclotus dysoni ambiguum .
Most cyclophorids share a similar reno-pericardial morphology ( Andrews & Little, 1972; Kasinathan, 1975). All except Murdochia pallidum ( Morton, 1952a) have a largely pallial kidney. This shift has been accompanied by a change in the topological relationships of features associated with the reno-pericardial complex such that the nephropore, afferent renal vessel and reno-pericardial canal all lie in a posterior position, with the nephropore opening to a posterior extension of the mantle cavity. The exact position of these features varies considerably among cyclophorid taxa ( Andrews & Little, 1972).
Although few accounts detail all aspects of cyclophorid nervous system morphology, those available converge on a consistent picture of a hypoathroid, right zygoneurous, left dialyneurous nervous system, lacking a labial commissure and possessing welldeveloped pedal cords ( Bouvier, 1887; Tielecke, 1940; Kasinathan, 1975). There are some conspicuous exceptions. For example, there are no reports of a small ganglion on the nerve from the left pleural ganglion. Given that osphradial innervation is primarily pleural among architaenioglossans ( Bouvier, 1887; Annandale & Sewell, 1921; Prashad, 1925; Berthold, 1991; Pon- der & Lindberg, 1997), the ganglion is undoubtedly an osphradial ganglion, despite loss of the osphradium. Bouvier (1887) and Kasinathan (1975) reported the presence of a single visceral ganglion and numerous cross-connections between the pedal cords in two species of Cyclophorus .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.